

Girdin is an intrinsic regulator of neuroblast chain migration in the rostral migratory stream of the postnatal brain
- PMID: 21632933
- PMCID: PMC5491307
- DOI: 10.1523/JNEUROSCI.1130-11.2011
Girdin is an intrinsic regulator of neuroblast chain migration in the rostral migratory stream of the postnatal brain
Abstract
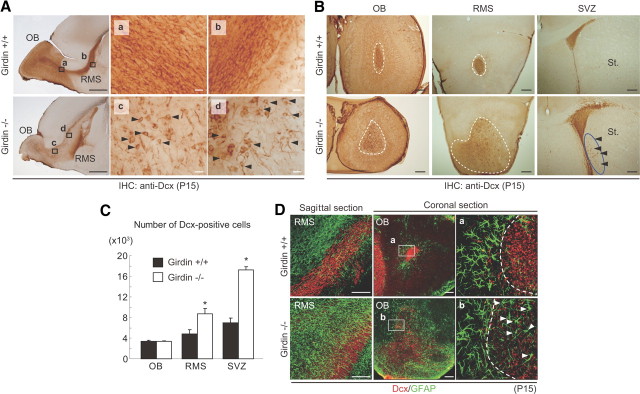
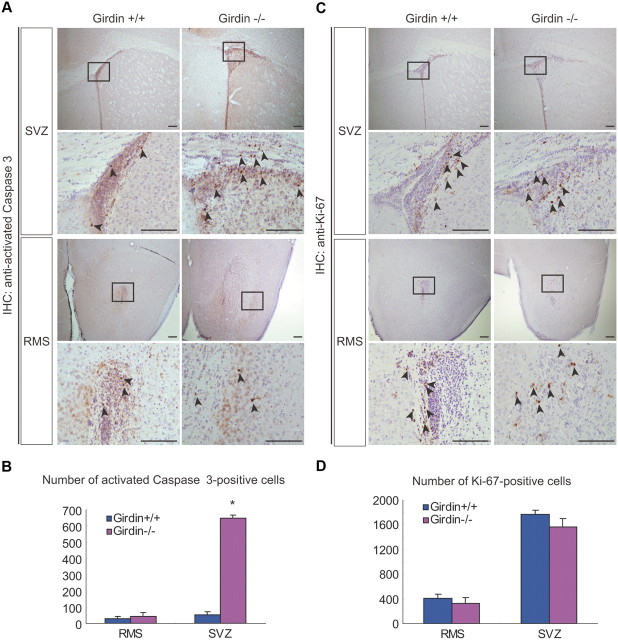
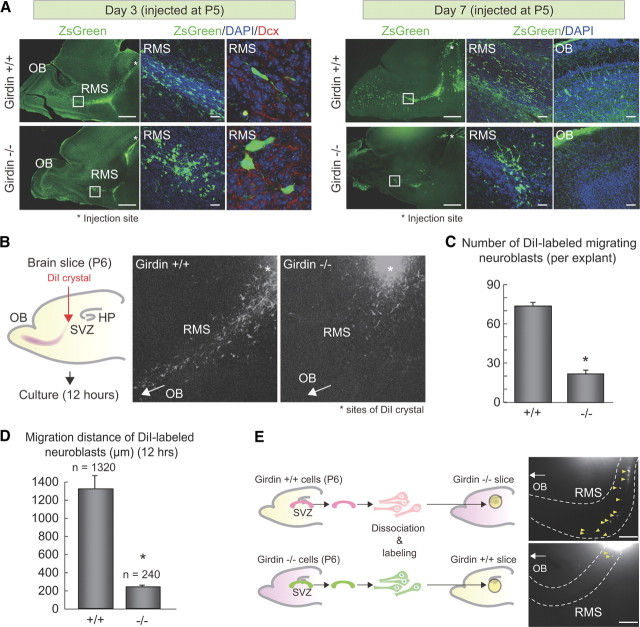
In postnatally developing and adult brains, interneurons of the olfactory bulb (OB) are continuously generated at the subventricular zone of the forebrain. The newborn neuroblasts migrate tangentially to the OB through a well defined pathway, the rostral migratory stream (RMS), where the neuroblasts undergo collective migration termed "chain migration." The cell-intrinsic regulatory mechanism of neuroblast chain migration, however, has not been uncovered. Here we show that mice lacking the actin-binding Akt substrate Girdin (a protein that interacts with Disrupted-In-Schizophrenia 1 to regulate neurogenesis in the dentate gyrus) have profound defects in neuroblast chain migration along the RMS. Analysis of two gene knock-in mice harboring Girdin mutants identified unique amino acid residues in Girdin's C-terminal domain that are responsible for the regulation of neuroblast chain migration but revealed no apparent requirement of Girdin phosphorylation by Akt. Electron microscopic analyses demonstrated the involvement of Girdin in neuroblast cell-cell interactions. These findings suggest that Girdin is an important intrinsic factor that specifically governs neuroblast chain migration along the RMS.
Figures







References
-
- Anton ES, Ghashghaei HT, Weber JL, McCann C, Fischer TM, Cheung IM, Gassmann M, Messing A, Klein R, Schwab MH, Lloyd KC, Lai C. Receptor tyrosine kinase ErbB4 modulates neuroblast migration and placement in the adult forebrain. Nat Neurosci. 2004;7:1319–1328. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous