

A mechanism of Rap1-induced stabilization of endothelial cell--cell junctions
- PMID: 21633110
- PMCID: PMC3135476
- DOI: 10.1091/mbc.E11-02-0157
A mechanism of Rap1-induced stabilization of endothelial cell--cell junctions
Abstract
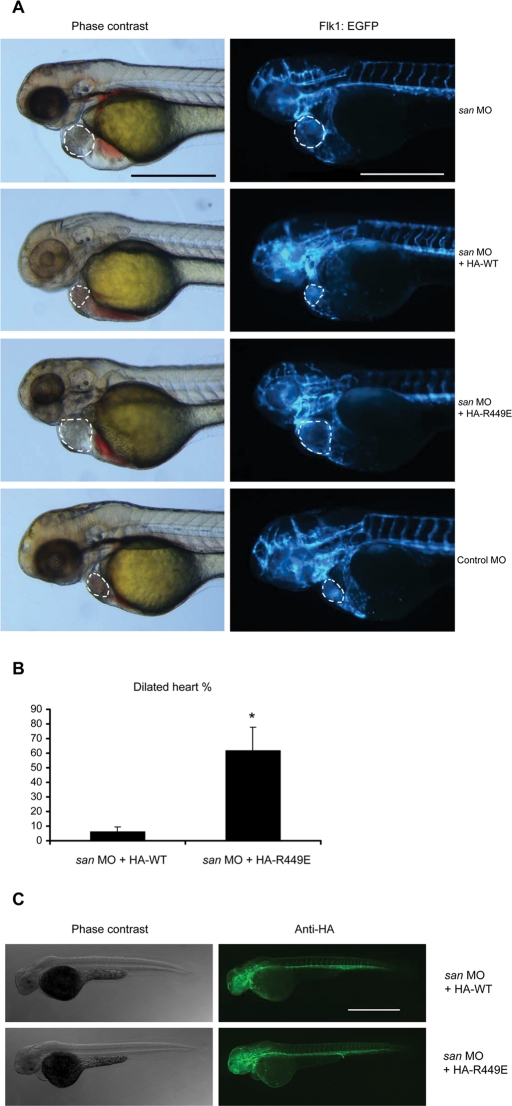
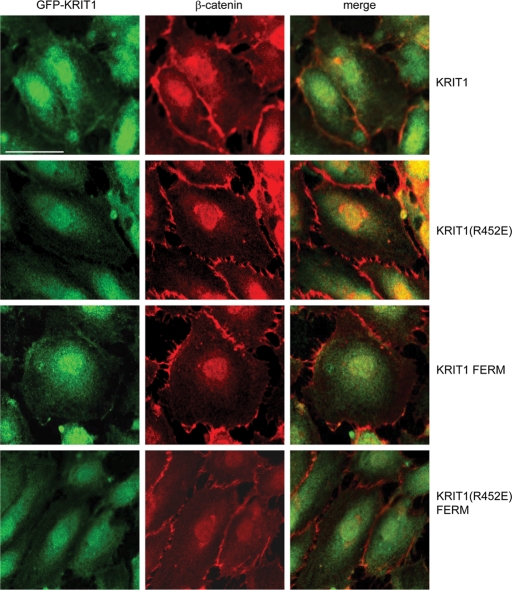
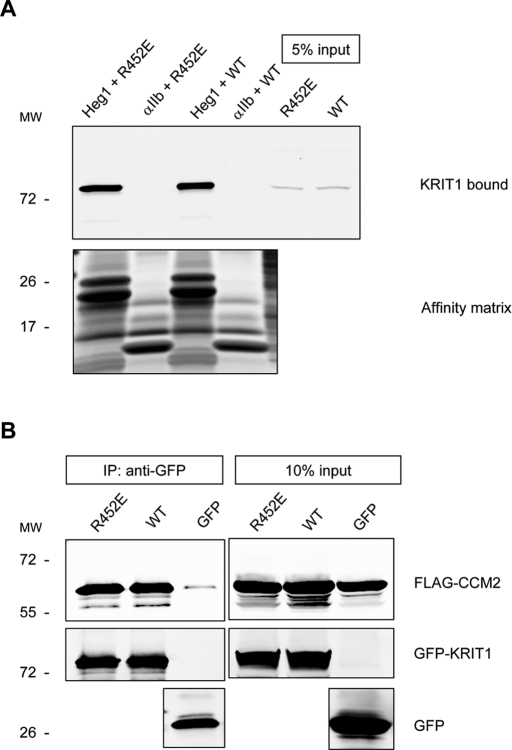
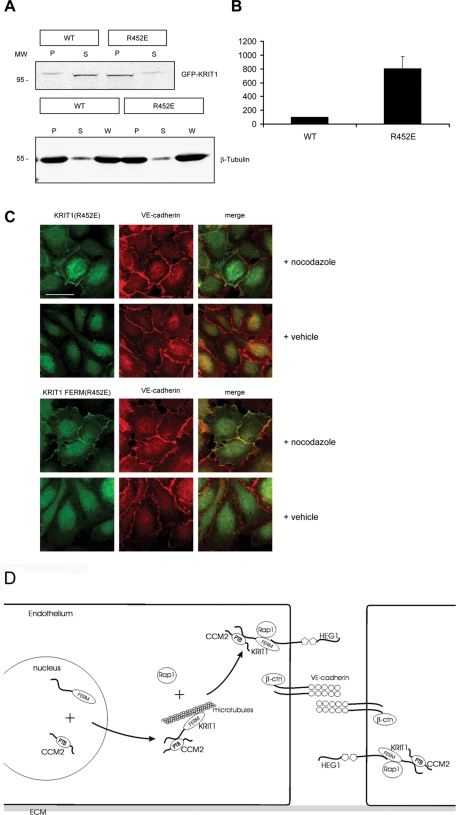
Activation of Rap1 small GTPases stabilizes cell--cell junctions, and this activity requires Krev Interaction Trapped gene 1 (KRIT1). Loss of KRIT1 disrupts cardiovascular development and causes autosomal dominant familial cerebral cavernous malformations. Here we report that native KRIT1 protein binds the effector loop of Rap1A but not H-Ras in a GTP-dependent manner, establishing that it is an authentic Rap1-specific effector. By modeling the KRIT1-Rap1 interface we designed a well-folded KRIT1 mutant that exhibited a ~40-fold-reduced affinity for Rap1A and maintained other KRIT1-binding functions. Direct binding of KRIT1 to Rap1 stabilized endothelial cell-cell junctions in vitro and was required for cardiovascular development in vivo. Mechanistically, Rap1 binding released KRIT1 from microtubules, enabling it to locate to cell--cell junctions, where it suppressed Rho kinase signaling and stabilized the junctions. These studies establish that the direct physical interaction of Rap1 with KRIT1 enables the translocation of microtubule-sequestered KRIT1 to junctions, thereby supporting junctional integrity and cardiovascular development.
Figures
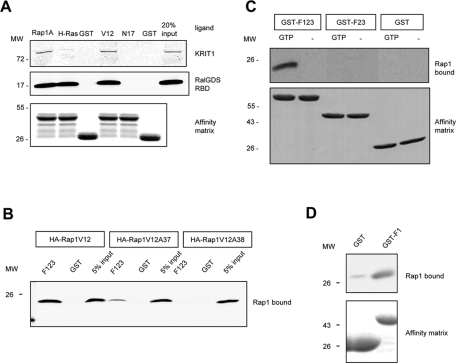
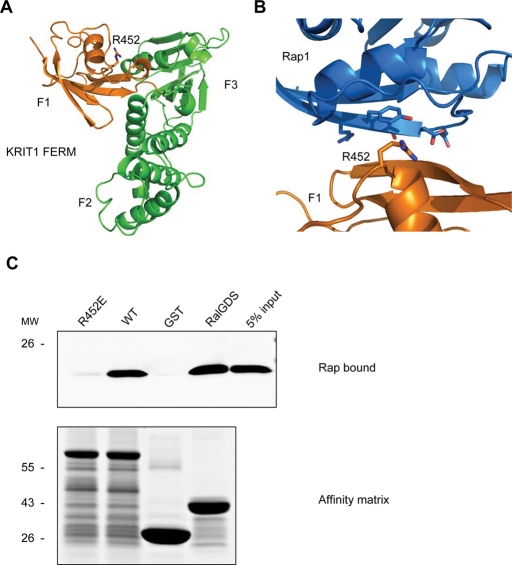
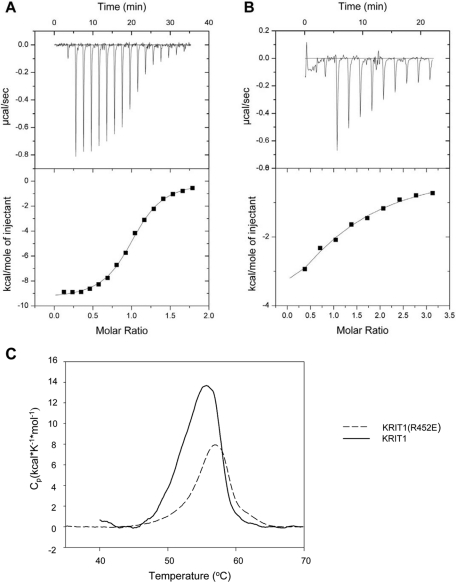
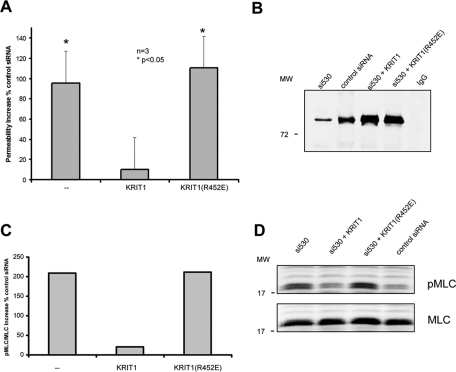
References
-
- Bos JL. Linking Rap to cell adhesion. Curr Opin Cell Biol. 2005;17:123–128. - PubMed
-
- Cullere X, Shaw SK, Andersson L, Hirahashi J, Luscinskas FW, Mayadas TN. Regulation of vascular endothelial barrier function by Epac, a cAMP-activated exchange factor for Rap GTPase. Blood. 2005;105:1950–1955. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
