

Bidirectional plasticity in the primate inferior olive induced by chronic ethanol intoxication and sustained abstinence
- PMID: 21642533
- PMCID: PMC3121872
- DOI: 10.1073/pnas.1017079108
Bidirectional plasticity in the primate inferior olive induced by chronic ethanol intoxication and sustained abstinence
Abstract
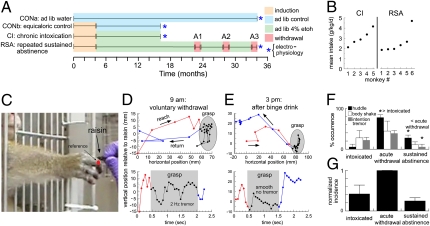
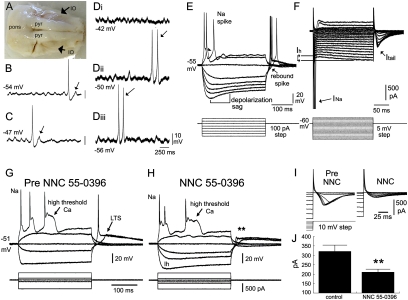
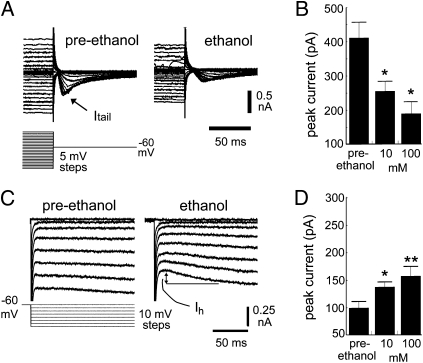
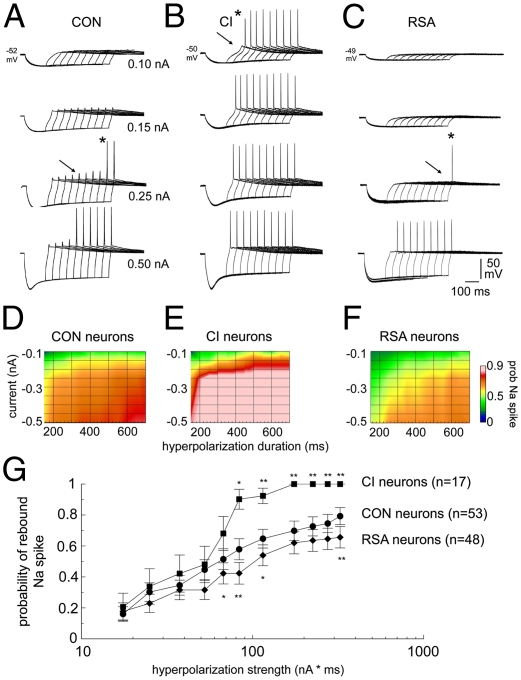
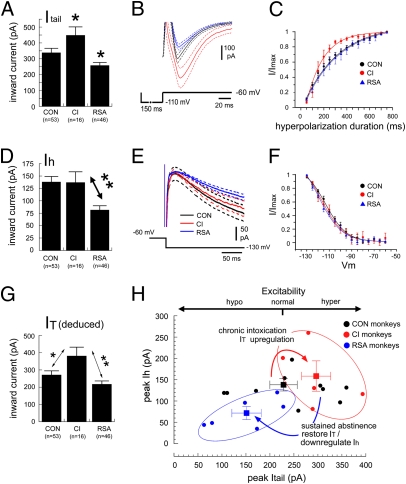
The brain adapts to chronic ethanol intoxication by altering synaptic and ion-channel function to increase excitability, a homeostatic counterbalance to inhibition by alcohol. Delirium tremens occurs when those adaptations are unmasked during withdrawal, but little is known about whether the primate brain returns to normal with repeated bouts of ethanol abuse and abstinence. Here, we show a form of bidirectional plasticity of pacemaking currents induced by chronic heavy drinking within the inferior olive of cynomolgus monkeys. Intracellular recordings of inferior olive neurons demonstrated that ethanol inhibited the tail current triggered by release from hyperpolarization (I(tail)). Both the slow deactivation of hyperpolarization-activated cyclic nucleotide-gated channels conducting the hyperpolarization-activated inward current and the activation of Ca(v)3.1 channels conducting the T-type calcium current (I(T)) contributed to I(tail), but ethanol inhibited only the I(T) component of I(tail). Recordings of inferior olive neurons obtained from chronically intoxicated monkeys revealed a significant up-regulation in I(tail) that was induced by 1 y of daily ethanol self-administration. The up-regulation was caused by a specific increase in I(T) which (i) greatly increased neurons' susceptibility for rebound excitation following hyperpolarization and (ii) may have accounted for intention tremors observed during ethanol withdrawal. In another set of monkeys, sustained abstinence produced the opposite effects: (i) a reduction in rebound excitability and (ii) a down-regulation of I(tail) caused by the down-regulation of both the hyperpolarization-activated inward current and I(T). Bidirectional plasticity of two hyperpolarization-sensitive currents following chronic ethanol abuse and abstinence may underlie persistent brain dysfunction in primates and be a target for therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Harris RA. Ethanol actions on multiple ion channels: Which are important? Alcohol Clin Exp Res. 1999;23:1563–1570. - PubMed
-
- Yang L, Long C, Evans MS, Faingold C. Ethanol withdrawal results in aberrant membrane properties and synaptic responses in periaqueductal gray neurons associated with seizure susceptibility. Brain Res. 2002;957:99–108. - PubMed
-
- Grant KA, Bennett AJ. Advances in nonhuman primate alcohol abuse and alcoholism research. Pharmacol Ther. 2003;100:235–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
