

UNC119 is required for G protein trafficking in sensory neurons
- PMID: 21642972
- PMCID: PMC3178889
- DOI: 10.1038/nn.2835
UNC119 is required for G protein trafficking in sensory neurons
Abstract
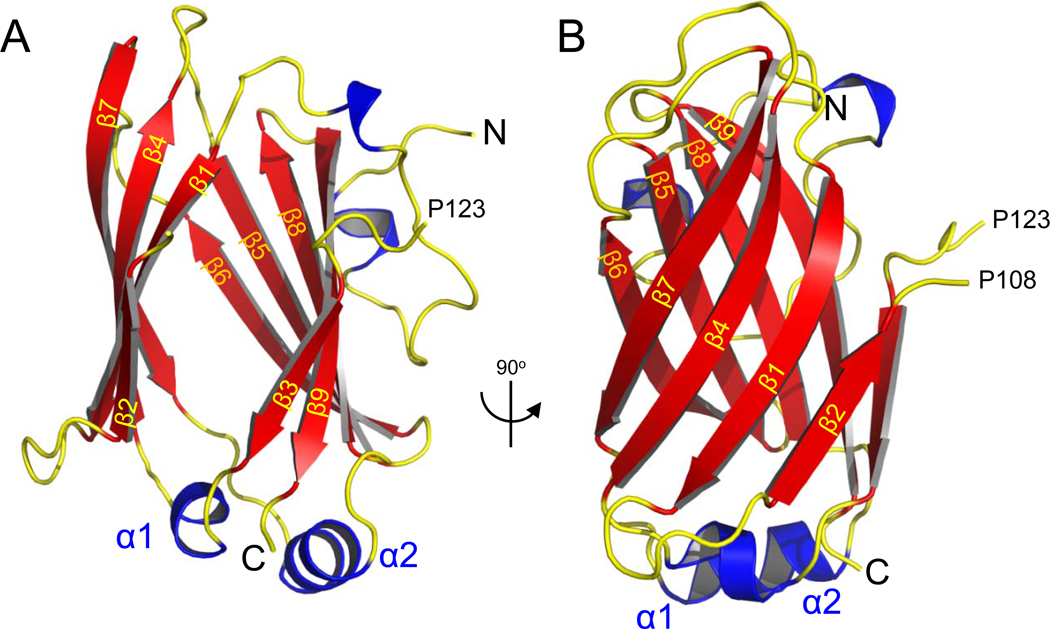
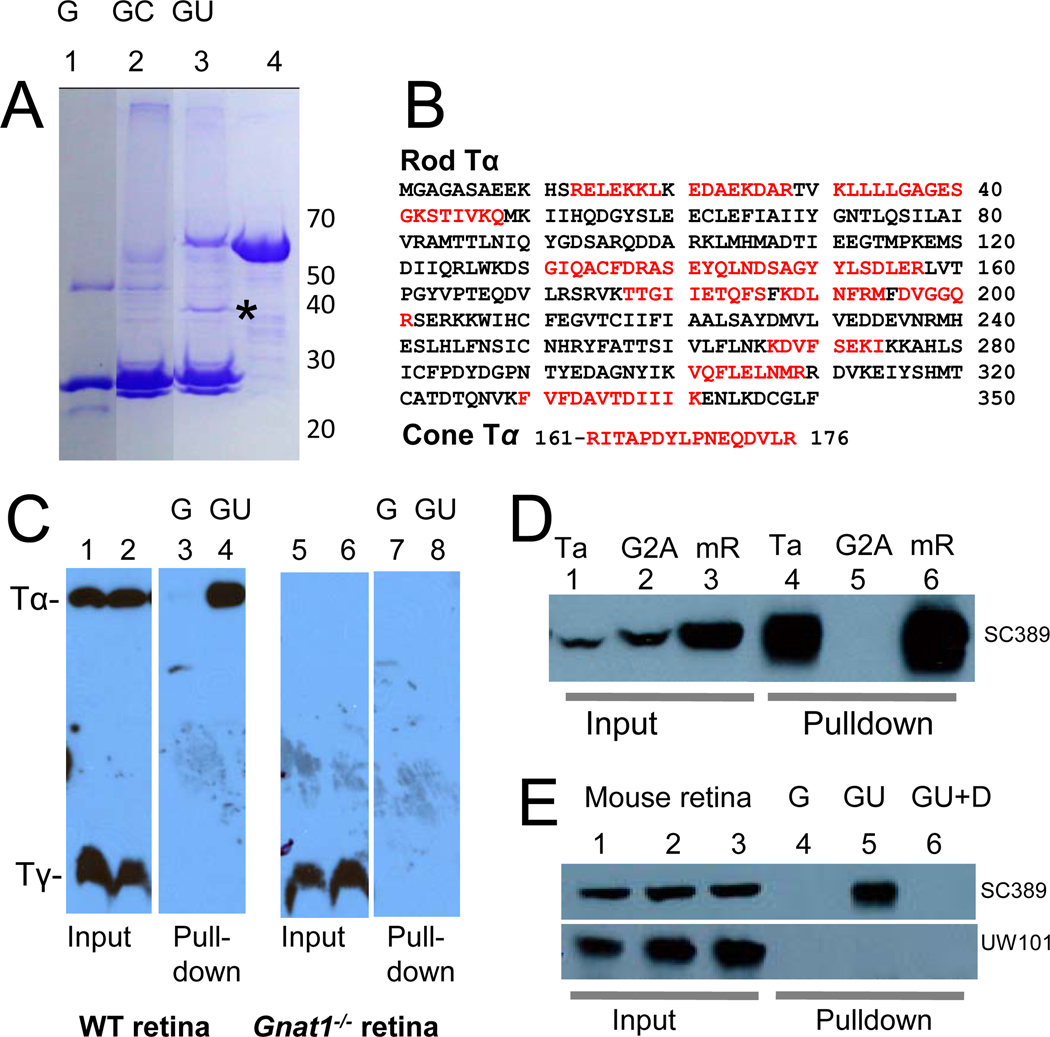
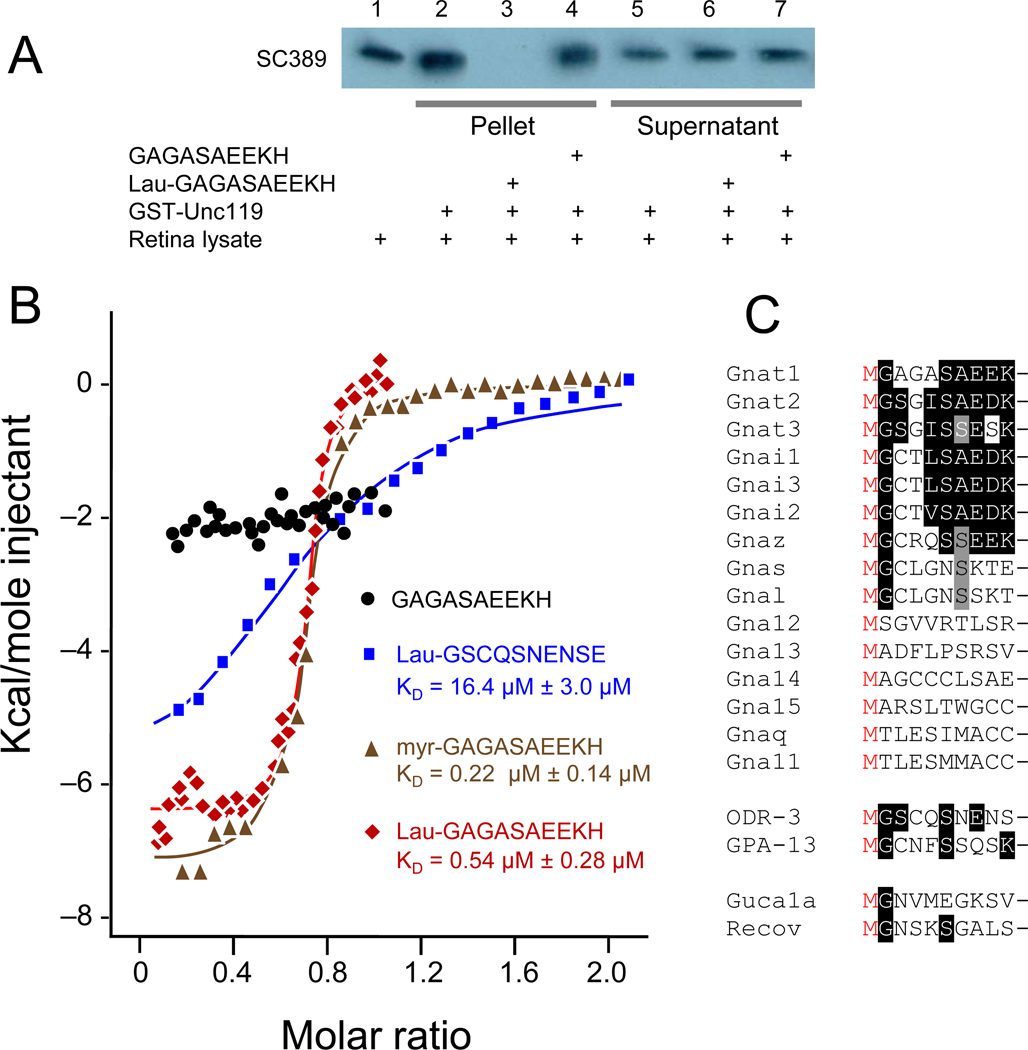
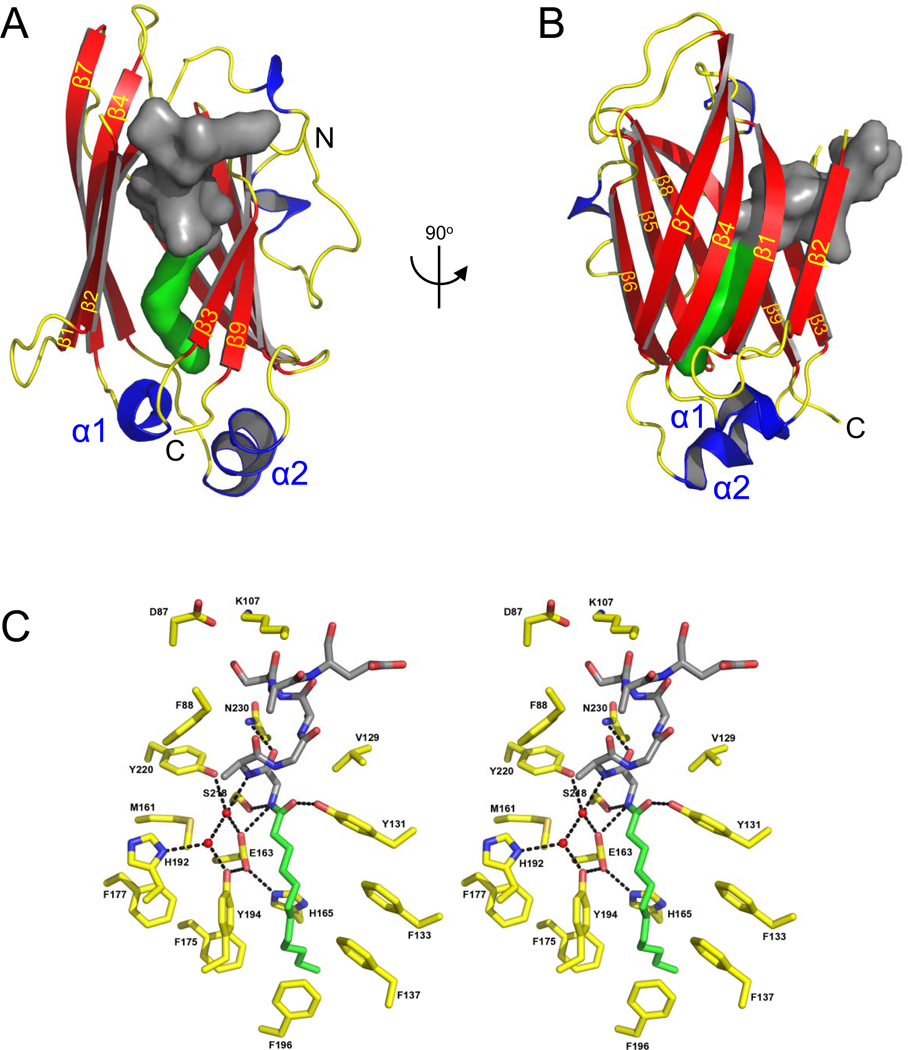
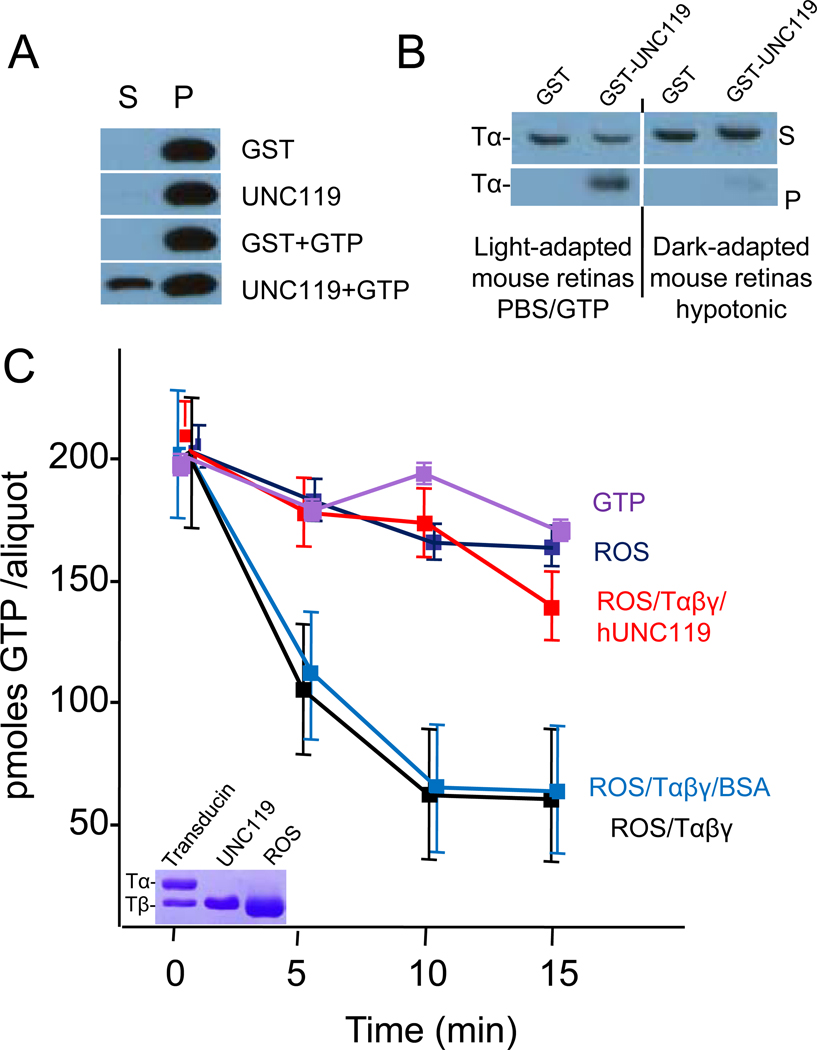
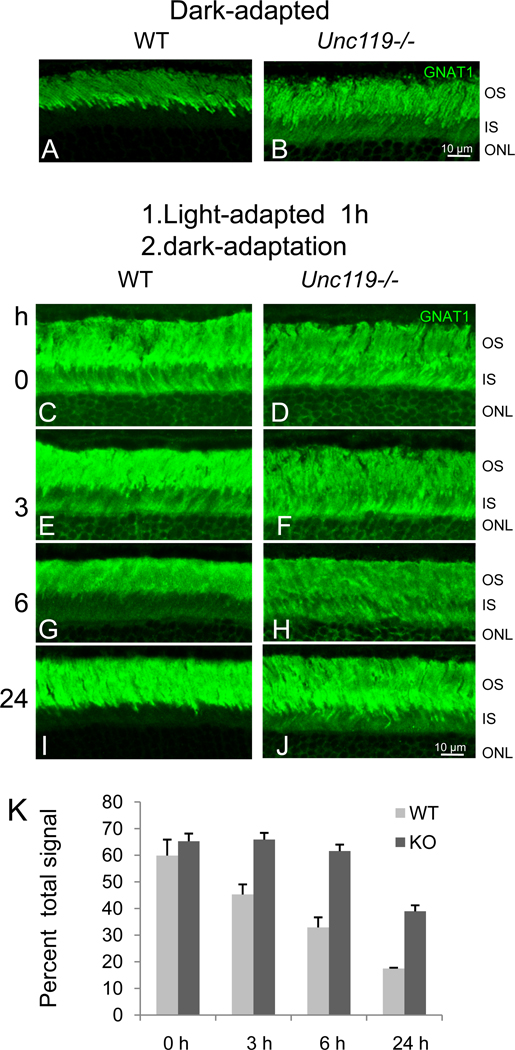
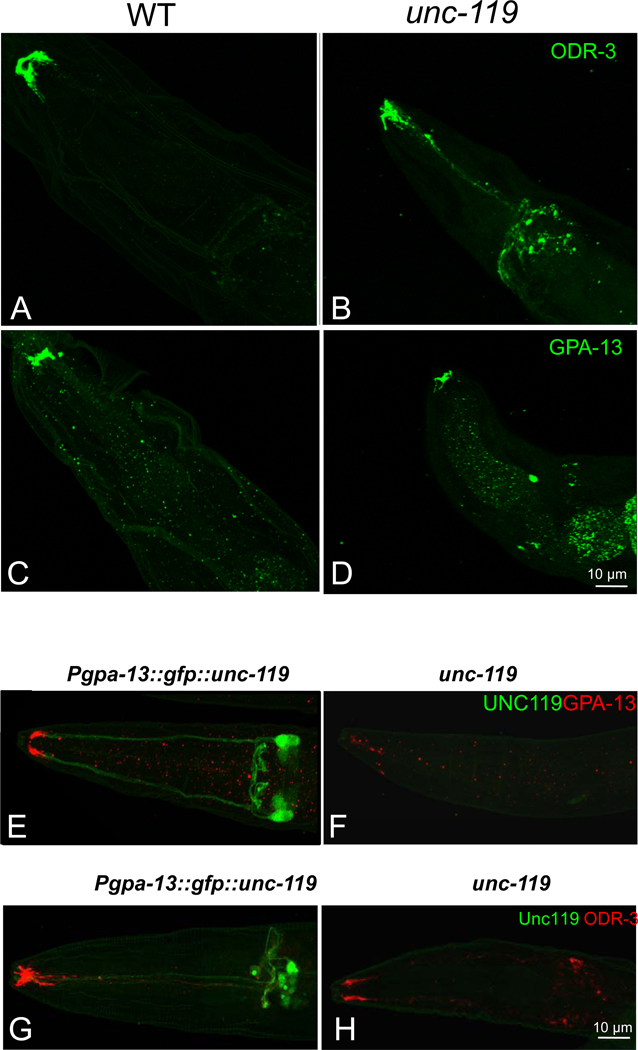
UNC119 is widely expressed among vertebrates and other phyla. We found that UNC119 recognized the acylated N terminus of the rod photoreceptor transducin α (Tα) subunit and Caenorhabditis elegans G proteins ODR-3 and GPA-13. The crystal structure of human UNC119 at 1.95-Å resolution revealed an immunoglobulin-like β-sandwich fold. Pulldowns and isothermal titration calorimetry revealed a tight interaction between UNC119 and acylated Gα peptides. The structure of co-crystals of UNC119 with an acylated Tα N-terminal peptide at 2.0 Å revealed that the lipid chain is buried deeply into UNC119's hydrophobic cavity. UNC119 bound Tα-GTP, inhibiting its GTPase activity, thereby providing a stable UNC119-Tα-GTP complex capable of diffusing from the inner segment back to the outer segment after light-induced translocation. UNC119 deletion in both mouse and C. elegans led to G protein mislocalization. Thus, UNC119 is a Gα subunit cofactor essential for G protein trafficking in sensory cilia.
Figures
Comment in
-
Vision and olfaction say UNC-le to G proteins.Nat Neurosci. 2011 Jun 27;14(7):805-6. doi: 10.1038/nn.2863. Nat Neurosci. 2011. PMID: 21709674 No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
