

Neuronal basis of sequential foraging decisions in a patchy environment
- PMID: 21642973
- PMCID: PMC3553855
- DOI: 10.1038/nn.2856
Neuronal basis of sequential foraging decisions in a patchy environment
Abstract
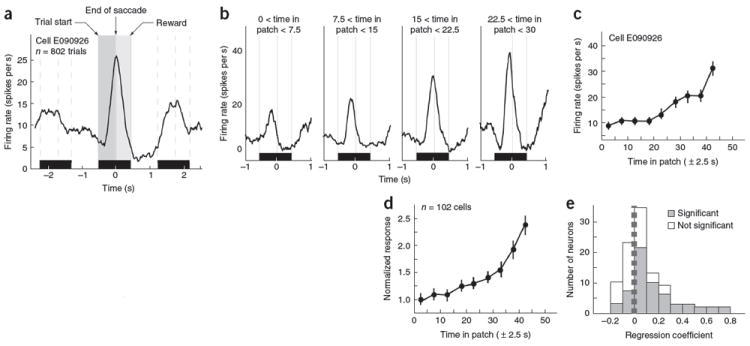
Deciding when to leave a depleting resource to exploit another is a fundamental problem for all decision makers. The neuronal mechanisms mediating patch-leaving decisions remain unknown. We found that neurons in primate (Macaca mulatta) dorsal anterior cingulate cortex, an area that is linked to reward monitoring and executive control, encode a decision variable signaling the relative value of leaving a depleting resource for a new one. Neurons fired during each sequential decision to stay in a patch and, for each travel time, these responses reached a fixed threshold for patch-leaving. Longer travel times reduced the gain of neural responses for choosing to stay in a patch and increased the firing rate threshold mandating patch-leaving. These modulations more closely matched behavioral decisions than any single task variable. These findings portend an understanding of the neural basis of foraging decisions and endorse the unification of theoretical and experimental work in ecology and neuroscience.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




References
-
- Prins HHT. Ecology and Behavior of the African Buffalo: Social Inequality and Decision Making. Chapman and Hall; London: 1996.
-
- Charnov EL. Optimal foraging, the marginal value theorem. Theor Popul Biol. 1976;9:129–136. - PubMed
-
- Stephens DW, Krebs JR. Foraging Theory. Princeton University Press; Princeton, NJ: 1986.
-
- Thompson D, Fedak MA. How long should a dive last? A simple model of foraging decisions by breath-hold divers in a patchy environment. Anim Behav. 2001;61:287–296.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
