

High-fat feeding promotes obesity via insulin receptor/PI3K-dependent inhibition of SF-1 VMH neurons
- PMID: 21642975
- PMCID: PMC3371271
- DOI: 10.1038/nn.2847
High-fat feeding promotes obesity via insulin receptor/PI3K-dependent inhibition of SF-1 VMH neurons
Abstract
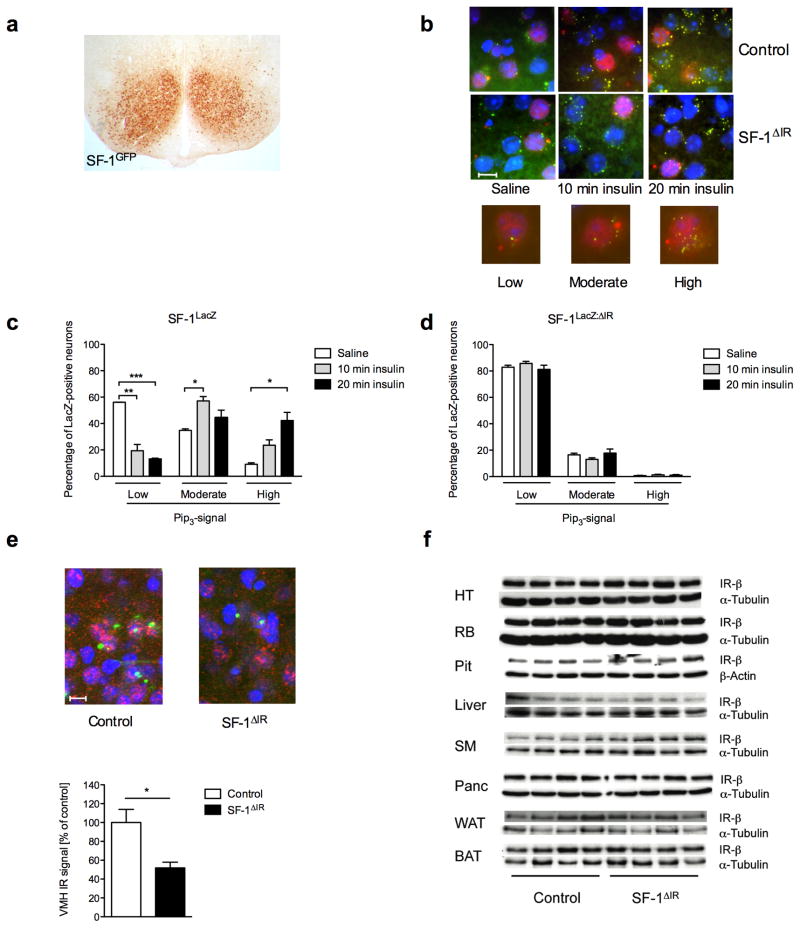
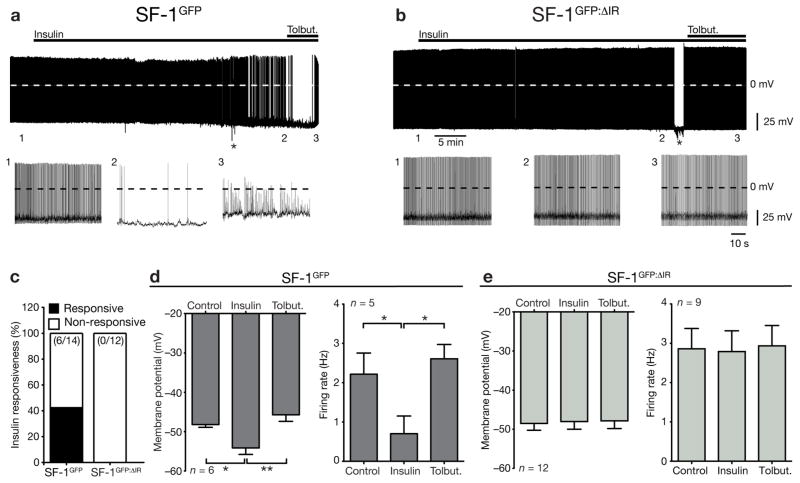
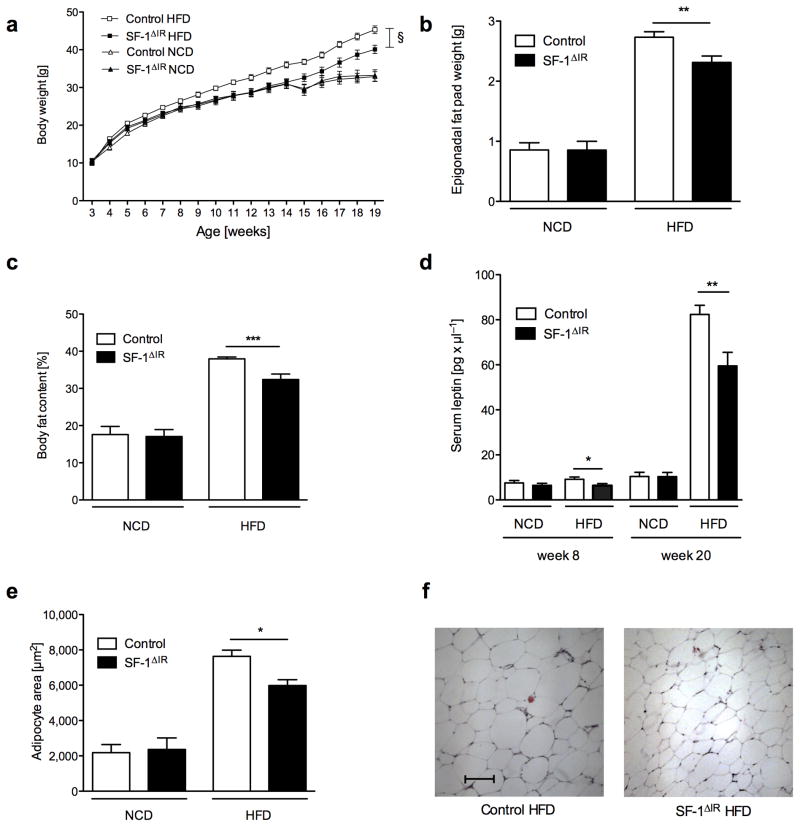
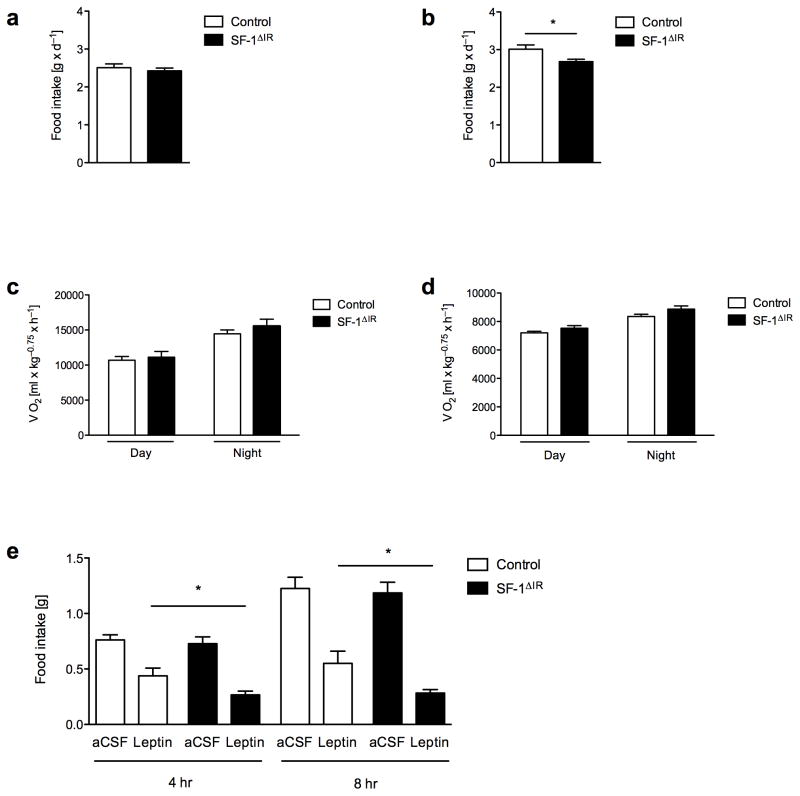
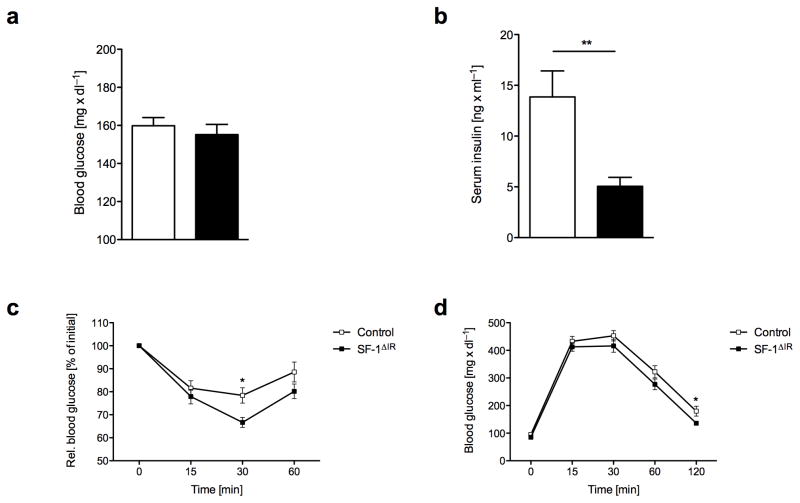
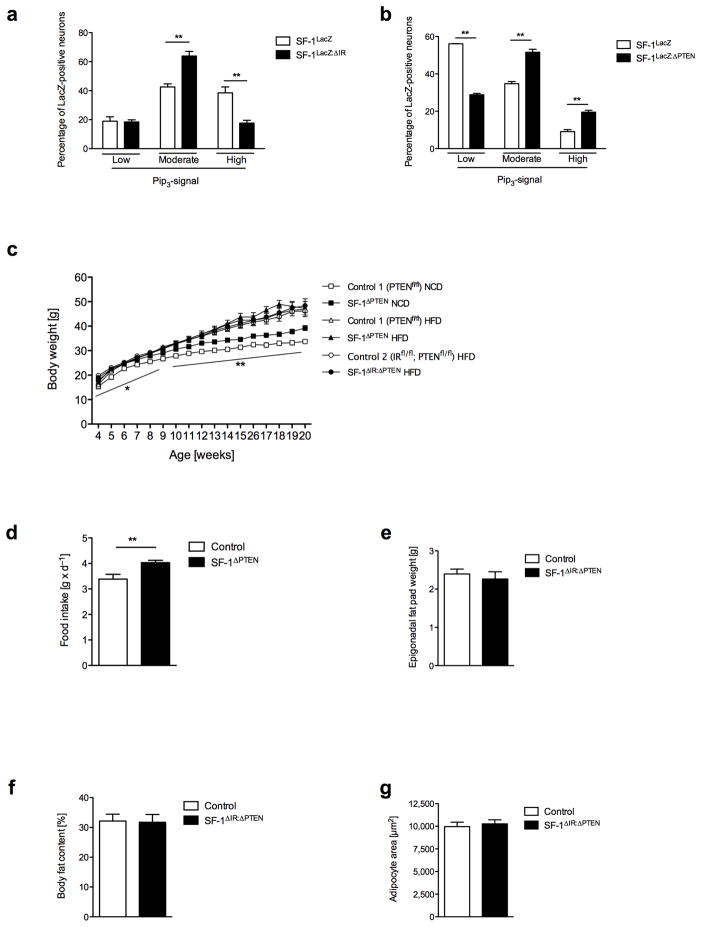
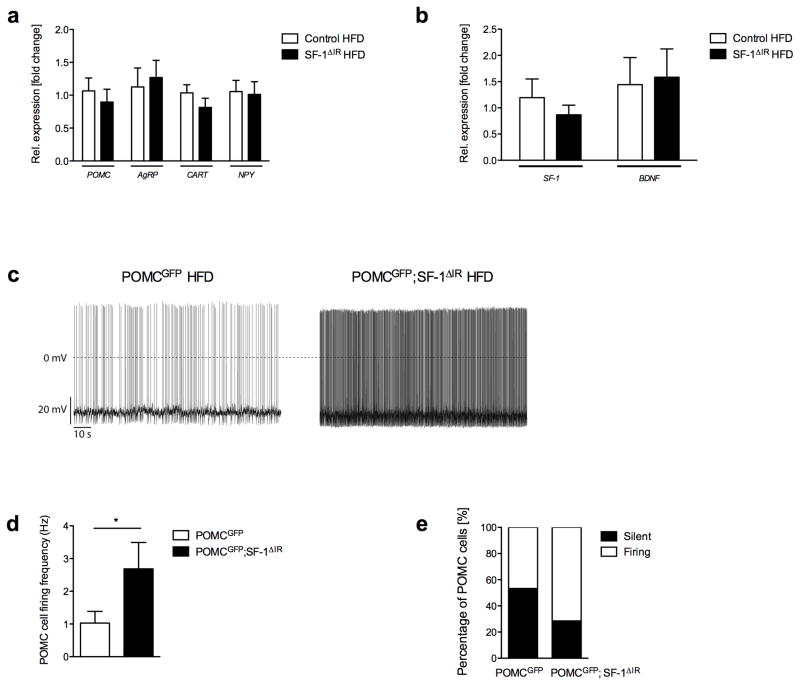
Steroidogenic factor 1 (SF-1)-expressing neurons of the ventromedial hypothalamus (VMH) control energy homeostasis, but the role of insulin action in these cells remains undefined. We show that insulin activates phosphatidylinositol-3-OH kinase (PI3K) signaling in SF-1 neurons and reduces firing frequency in these cells through activation of K(ATP) channels. These effects were abrogated in mice with insulin receptor deficiency restricted to SF-1 neurons (SF-1(ΔIR) mice). Whereas body weight and glucose homeostasis remained the same in SF-1(ΔIR) mice as in controls under a normal chow diet, they were protected from diet-induced leptin resistance, weight gain, adiposity and impaired glucose tolerance. High-fat feeding activated PI3K signaling in SF-1 neurons of control mice, and this response was attenuated in the VMH of SF-1(ΔIR) mice. Mimicking diet-induced overactivation of PI3K signaling by disruption of the phosphatidylinositol-3,4,5-trisphosphate phosphatase PTEN led to increased body weight and hyperphagia under a normal chow diet. Collectively, our experiments reveal that high-fat diet-induced, insulin-dependent PI3K activation in VMH neurons contributes to obesity development.
Figures
Comment in
-
Cajal revisited: does the VMH make us fat?Nat Neurosci. 2011 Jun 27;14(7):806-8. doi: 10.1038/nn.2867. Nat Neurosci. 2011. PMID: 21709675 No abstract available.
References
-
- Ikeda Y, Luo X, Abbud R, Nilson JH, Parker KL. The nuclear receptor steroidogenic factor 1 is essential for the formation of the ventromedial hypothalamic nucleus. Mol Endocrinol. 1995;9:478–486. - PubMed
-
- Sadovsky Y, et al. Mice deficient in the orphan receptor steroidogenic factor 1 lack adrenal glands and gonads but express P450 side-chain-cleavage enzyme in the placenta and have normal embryonic serum levels of corticosteroids. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:10939–10943. - PMC - PubMed
-
- Shinoda K, et al. Developmental defects of the ventromedial hypothalamic nucleus and pituitary gonadotroph in the Ftz-F1 disrupted mice. Dev Dyn. 1995;204:22–29. - PubMed
-
- Majdic G, et al. Knockout mice lacking steroidogenic factor 1 are a novel genetic model of hypothalamic obesity. Endocrinology. 2002;143:607–614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K08 DK068069-01A2/DK/NIDDK NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
- RL1 DK081185/DK/NIDDK NIH HHS/United States
- R01 DK080000/DK/NIDDK NIH HHS/United States
- K01DK087780/DK/NIDDK NIH HHS/United States
- OD006850/OD/NIH HHS/United States
- RL1DK081185/DK/NIDDK NIH HHS/United States
- UL1RR024923/RR/NCRR NIH HHS/United States
- DK080000/DK/NIDDK NIH HHS/United States
- DP1 OD006850/OD/NIH HHS/United States
- R01DK53301/DK/NIDDK NIH HHS/United States
- R01 DK071051/DK/NIDDK NIH HHS/United States
- R01 DK053301/DK/NIDDK NIH HHS/United States
- P30 DK046200/DK/NIDDK NIH HHS/United States
- K08 DK068069/DK/NIDDK NIH HHS/United States
- K01 DK087780/DK/NIDDK NIH HHS/United States
- UL1 RR024923/RR/NCRR NIH HHS/United States
- PL1 DK081182/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
