

CD4 T cells play important roles in maintaining IL-17-producing γδ T-cell subsets in naive animals
- PMID: 21647171
- PMCID: PMC3170686
- DOI: 10.1038/icb.2011.50
CD4 T cells play important roles in maintaining IL-17-producing γδ T-cell subsets in naive animals
Abstract
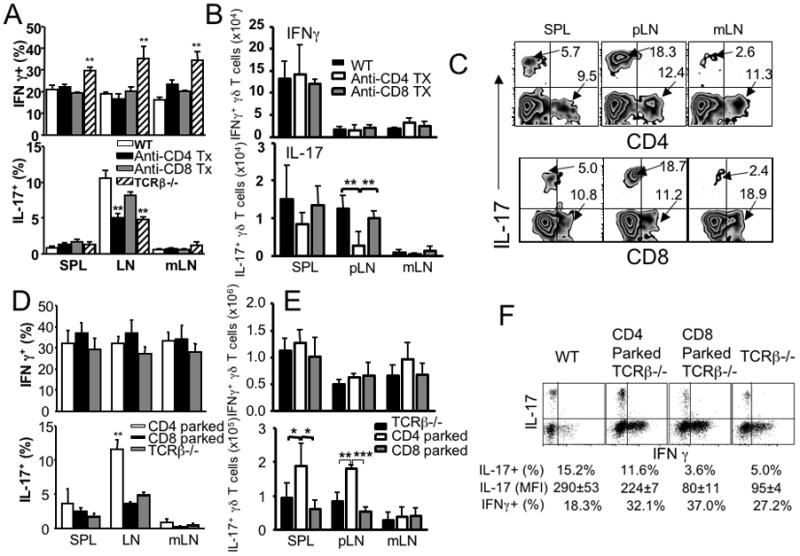
A proportional balance between αβ and γδ T-cell subsets in the periphery is exceedingly well maintained by a homeostatic mechanism. However, a cellular mechanism underlying the regulation remains undefined. We recently reported that a subset of developing γδ T cells spontaneously acquires interleukin (IL)-17-producing capacity even within naive animals through a transforming growth factor (TGF)β1-dependent mechanism, thus considered 'innate' IL-17-producing cells. Here, we report that γδ T cells generated within αβ T cell (or CD4 T cell)-deficient environments displayed altered cytokine profiles; particularly, 'innate' IL-17 expression was significantly impaired compared with those in wild-type mice. Impaired IL-17 production in γδ T cells was directly related to CD4 T-cell deficiency, because depletion of CD4 T cells in wild-type mice diminished and adoptive CD4 T-cell transfer into T-cell receptor β-/- mice restored IL-17 expression in γδ T cells. CD4 T cell-mediated IL-17 expression required TGFβ1. Moreover, Th17 but not Th1 or Th2 effector CD4 T cells were highly efficient in enhancing γδ T-cell IL-17 expression. Taken together, our results highlight a novel CD4 T cell-dependent mechanism that shapes the generation of IL-17+ γδ T cells in naive settings.
Figures






Comment in
-
γδ T cells are not alone.Immunol Cell Biol. 2012 Apr;90(4):370-1. doi: 10.1038/icb.2011.61. Epub 2011 Jul 12. Immunol Cell Biol. 2012. PMID: 21747406 No abstract available.
References
-
- Shibata K, Yamada H, Nakamura R, Sun X, Itsumi M, Yoshikai Y. Identification of CD25+ gamma delta T cells as fetal thymus-derived naturally occurring IL-17 producers. J Immunol. 2008;181(9):5940–7. - PubMed
-
- Haas JD, Gonzalez FH, Schmitz S, Chennupati V, Fohse L, Kremmer E, et al. CCR6 and NK1.1 distinguish between IL-17A and IFN-gamma-producing gammadelta effector T cells. Eur J Immunol. 2009;39(12):3488–97. - PubMed
-
- Kisielow J, Kopf M, Karjalainen K. SCART scavenger receptors identify a novel subset of adult gammadelta T cells. J Immunol. 2008;181(3):1710–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
