

Staphylococcus aureus regulates the expression and production of the staphylococcal superantigen-like secreted proteins in a Rot-dependent manner
- PMID: 21651625
- PMCID: PMC3217042
- DOI: 10.1111/j.1365-2958.2011.07720.x
Staphylococcus aureus regulates the expression and production of the staphylococcal superantigen-like secreted proteins in a Rot-dependent manner
Abstract
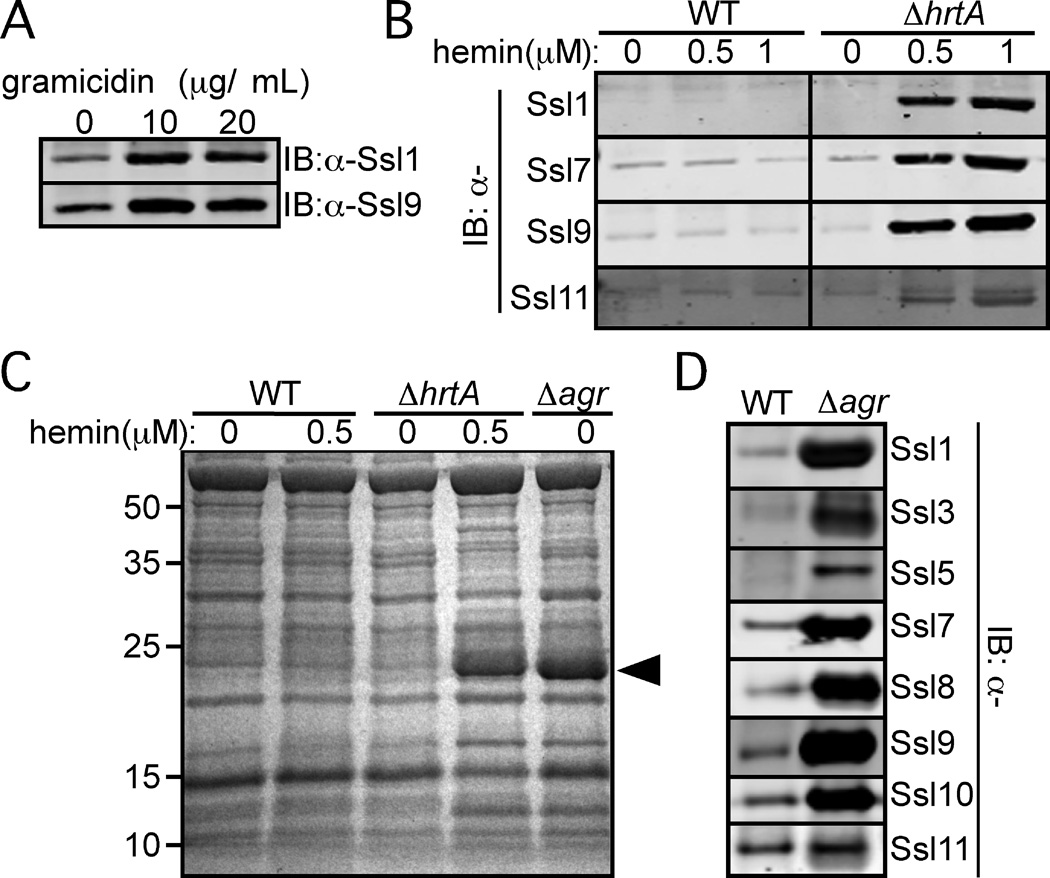
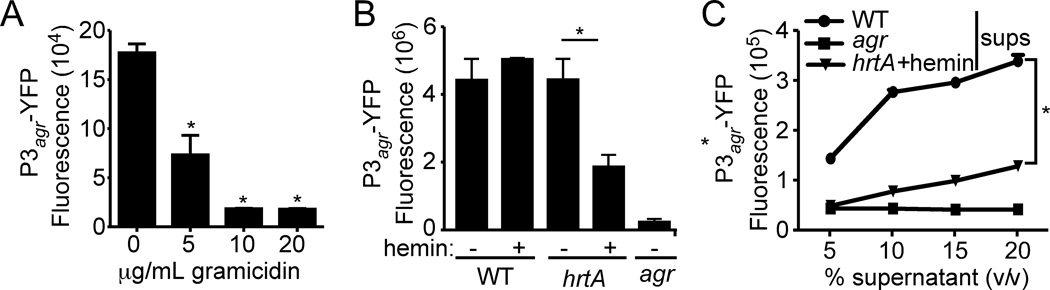
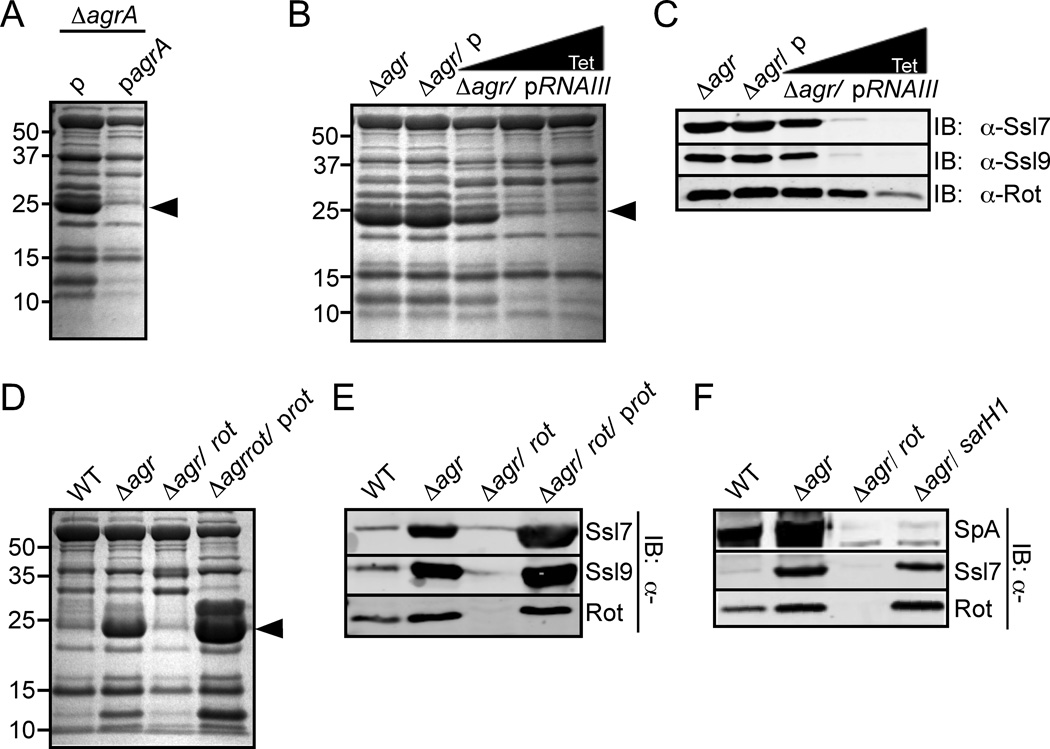
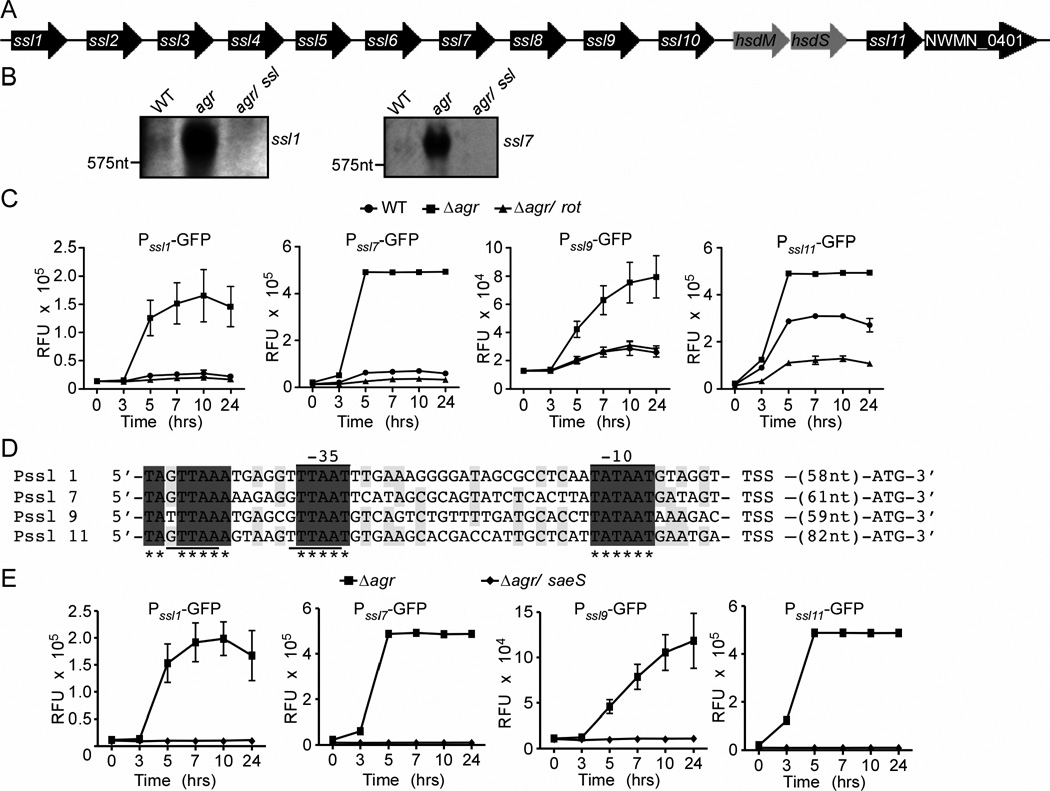
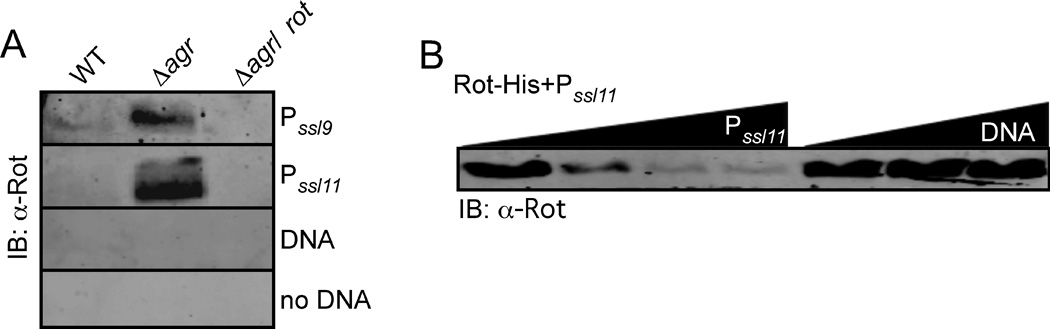
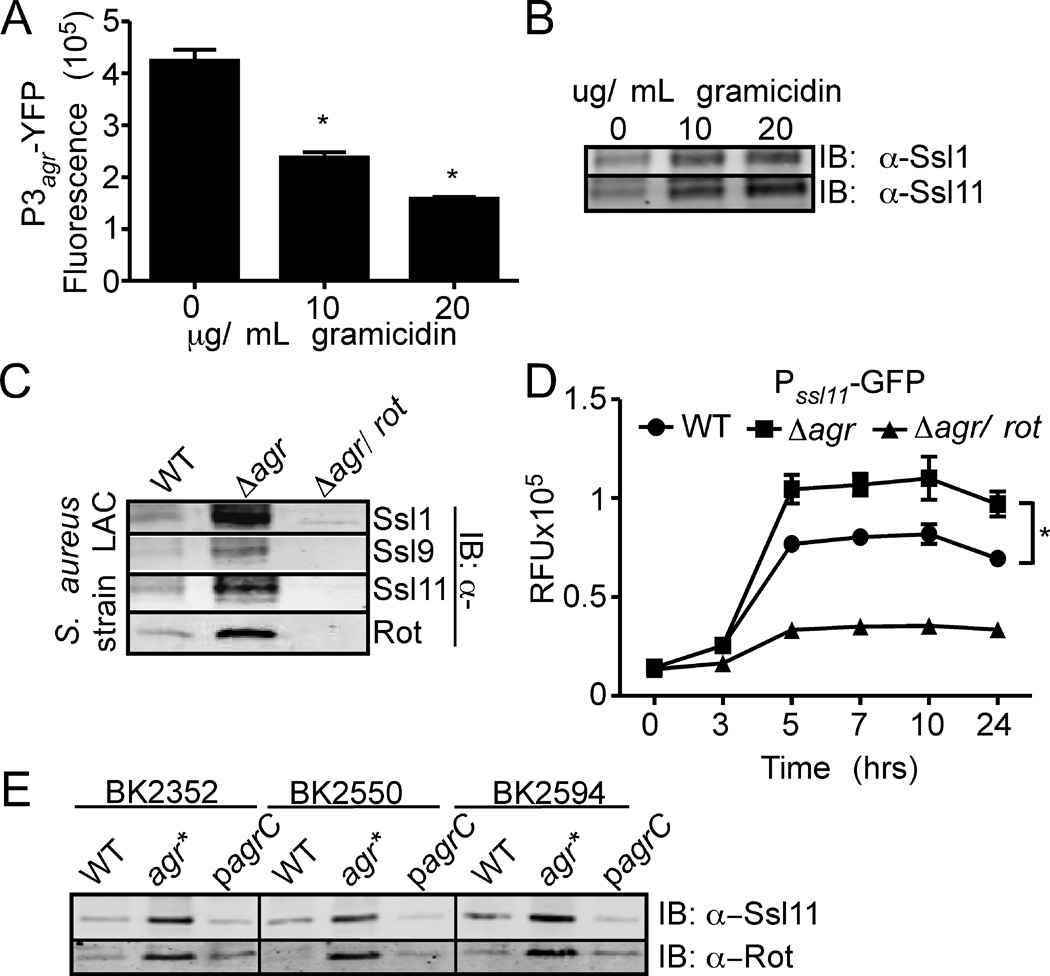
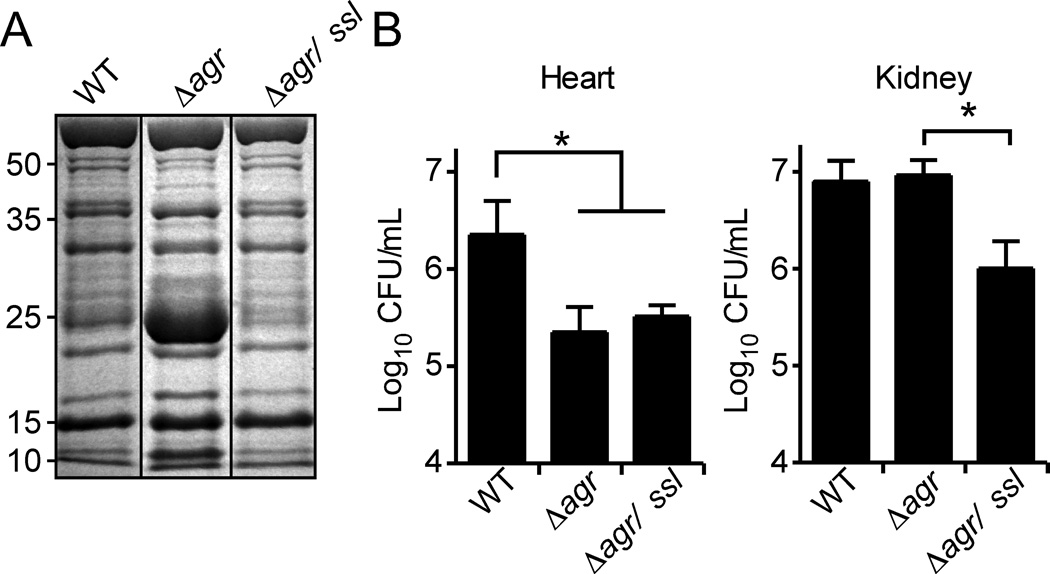
Staphylococcus aureus overproduces a subset of immunomodulatory proteins known as the staphylococcal superantigen-like proteins (Ssls) under conditions of pore-mediated membrane stress. In this study we demonstrate that overproduction of Ssls during membrane stress is due to the impaired activation of the two-component module of the quorum-sensing accessory gene regulator (Agr) system. Agr-dependent repression of ssl expression is indirect and mediated by the transcription factor repressor of toxins (Rot). Surprisingly, we observed that Rot directly interacts with and activates the ssl promoters. The role of Agr and Rot as regulators of ssl expression was observed across several clinically relevant strains, suggesting that overproduction of immunomodulatory proteins benefits agr-defective strains. In support of this notion, we demonstrate that Ssls contribute to the residual virulence of S. aureus lacking agr in a murine model of systemic infection. Altogether, these results suggest that S. aureus compensates for the inactivation of Agr by producing immunomodulatory exoproteins that could protect the bacterium from host-mediated clearance.
© 2011 Blackwell Publishing Ltd.
Figures
References
-
- Adhikari RP, Novick RP. Regulatory organization of the staphylococcal sae locus. Microbiology. 2008;154:949–959. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
