

A context-dependent combination of Wnt receptors controls axis elongation and leg development in a short germ insect
- PMID: 21652652
- PMCID: PMC3188592
- DOI: 10.1242/dev.063644
A context-dependent combination of Wnt receptors controls axis elongation and leg development in a short germ insect
Abstract
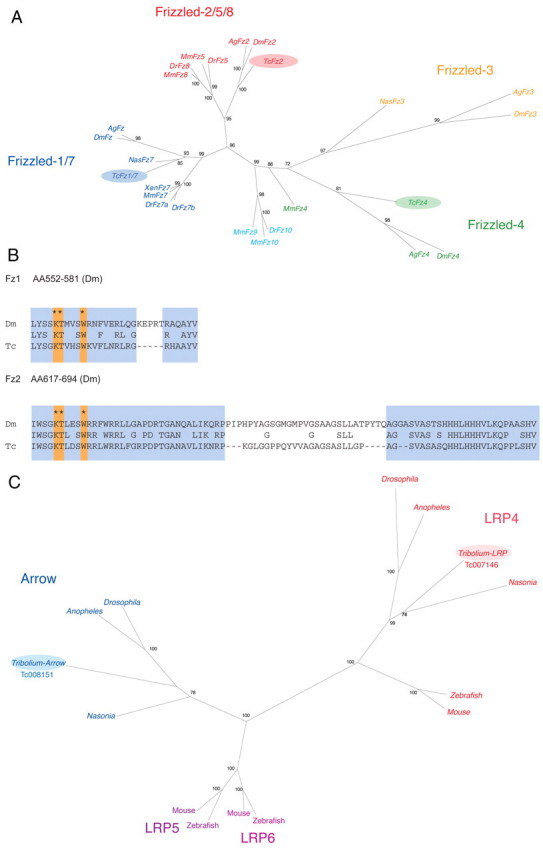
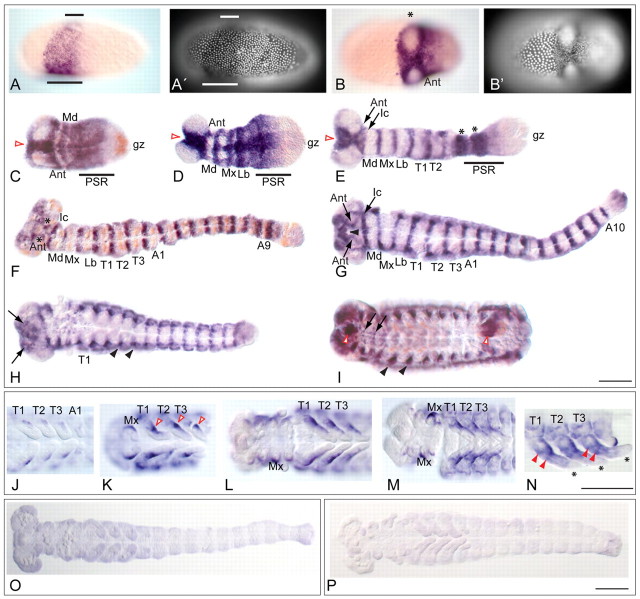
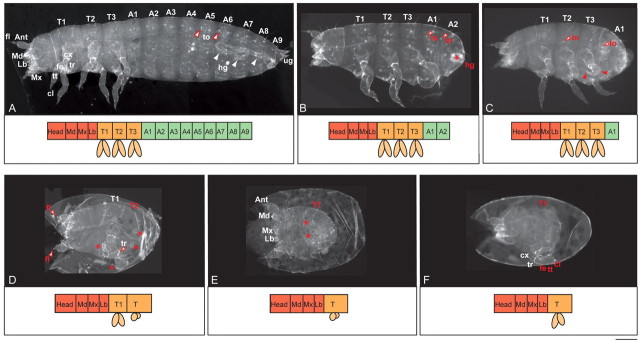
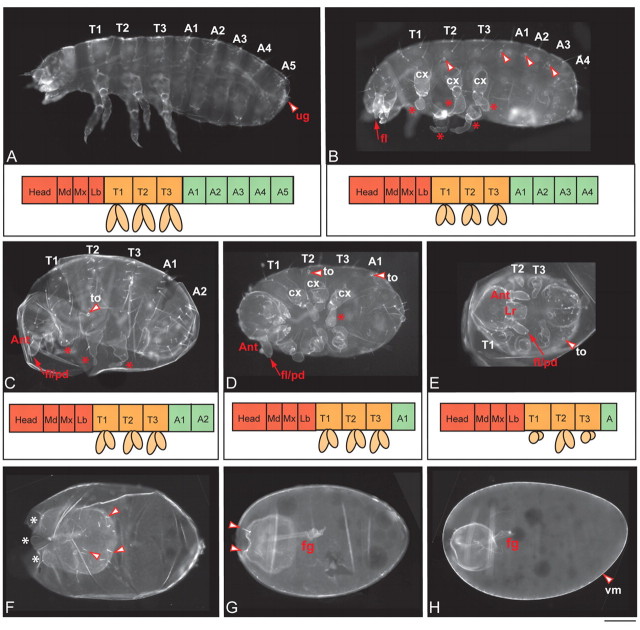
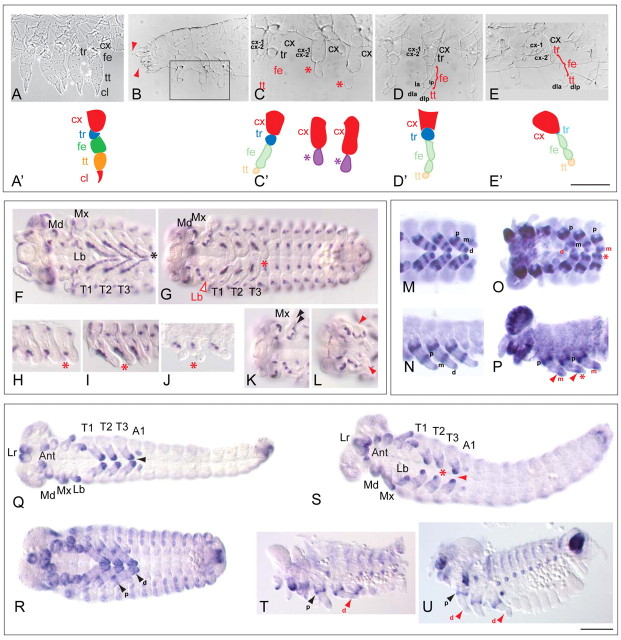
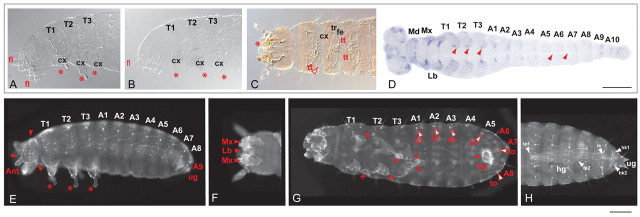
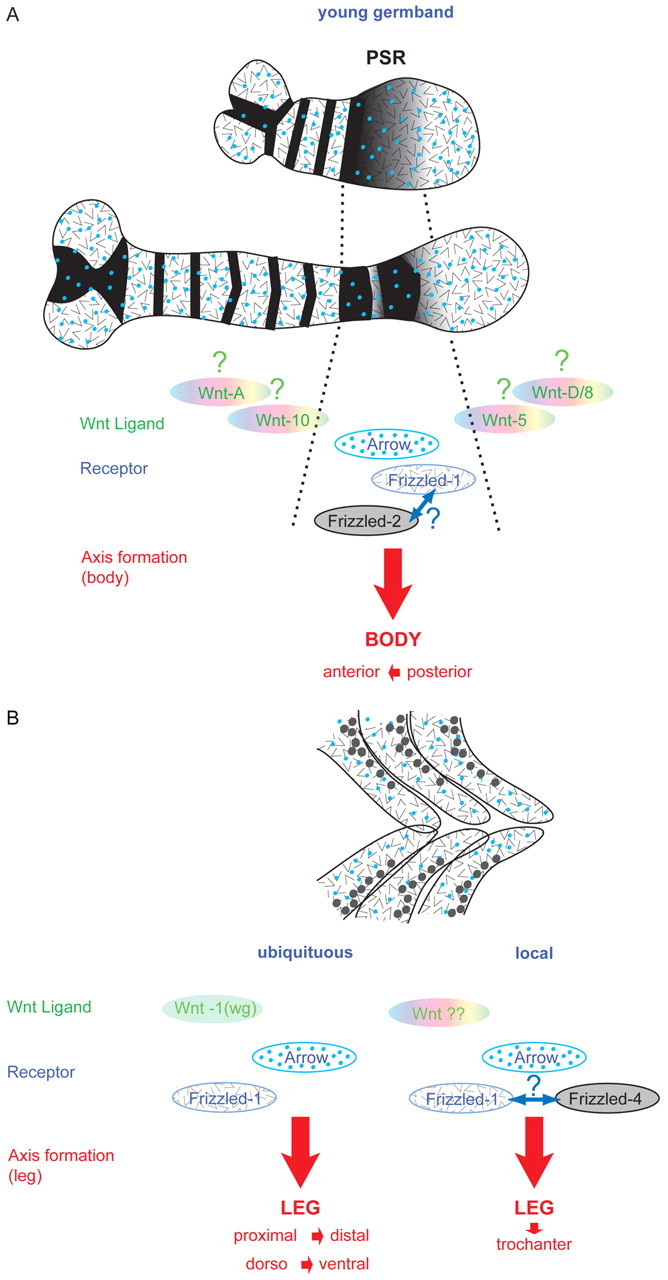
Short germ embryos elongate their primary body axis by consecutively adding segments from a posteriorly located growth zone. Wnt signalling is required for axis elongation in short germ arthropods, including Tribolium castaneum, but the precise functions of the different Wnt receptors involved in this process are unclear. We analysed the individual and combinatorial functions of the three Wnt receptors, Frizzled-1 (Tc-Fz1), Frizzled-2 (Tc-Fz2) and Frizzled-4 (Tc-Fz4), and their co-receptor Arrow (Tc-Arr) in the beetle Tribolium. Knockdown of gene function and expression analyses revealed that Frizzled-dependent Wnt signalling occurs anteriorly in the growth zone in the presegmental region (PSR). We show that simultaneous functional knockdown of the Wnt receptors Tc-fz1 and Tc-fz2 via RNAi resulted in collapse of the growth zone and impairment of embryonic axis elongation. Although posterior cells of the growth zone were not completely abolished, Wnt signalling within the PSR controls axial elongation at the level of pair-rule patterning, Wnt5 signalling and FGF signalling. These results identify the PSR in Tribolium as an integral tissue required for the axial elongation process, reminiscent of the presomitic mesoderm in vertebrates. Knockdown of Tc-fz1 alone interfered with the formation of the proximo-distal and the dorso-ventral axes during leg development, whereas no effect was observed with single Tc-fz2 or Tc-fz4 RNAi knockdowns. We identify Tc-Arr as an obligatory Wnt co-receptor for axis elongation, leg distalisation and segmentation. We discuss how Wnt signalling is regulated at the receptor and co-receptor levels in a dose-dependent fashion.
Figures

References
-
- Angers S., Salahpour A., Bouvier M. (2002). Dimerization: an emerging concept for G protein-coupled receptor ontogeny and function. Annu. Rev. Pharmacol. Toxicol. 42, 409-435 - PubMed
-
- Arnold S. J., Stappert J., Bauer A., Kispert A., Herrmann B. G., Kemler R. (2000). Brachyury is a target gene of the Wnt/beta-catenin signaling pathway. Mech. Dev. 91, 249-258 - PubMed
-
- Beermann A., Schröder R. (2008). Sites of Fgf signalling and perception during embryogenesis of the beetle Tribolium castaneum. Dev. Genes Evol. 218, 153-167 - PubMed
-
- Beermann A., Aranda M., Schröder R. (2004). The Sp8 zinc-finger transcription factor is involved in allometric growth of the limbs in the beetle Tribolium castaneum. Development 131, 733-742 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
