

Crosstalk between NOTCH and AKT signaling during murine megakaryocyte lineage specification
- PMID: 21653327
- PMCID: PMC3152494
- DOI: 10.1182/blood-2011-01-328567
Crosstalk between NOTCH and AKT signaling during murine megakaryocyte lineage specification
Abstract
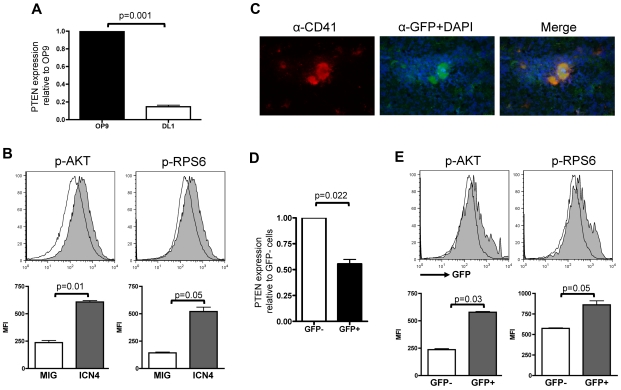
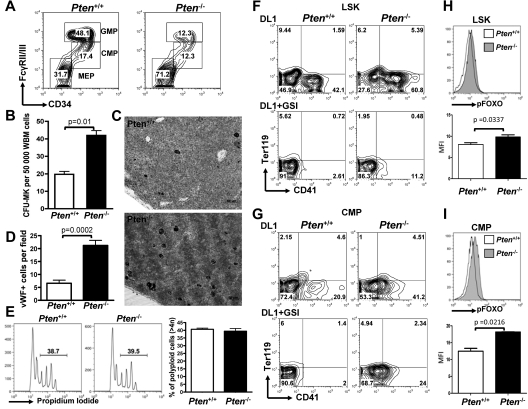
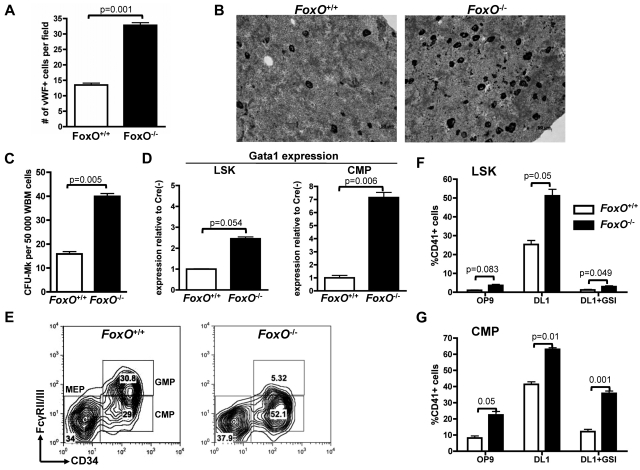
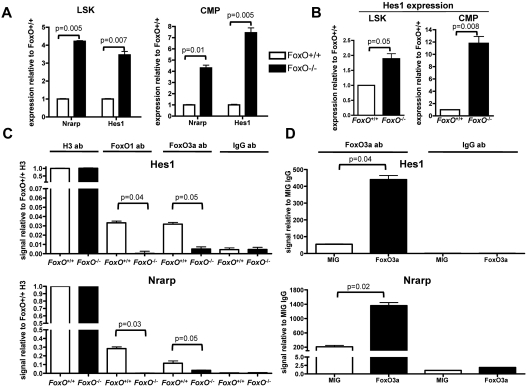
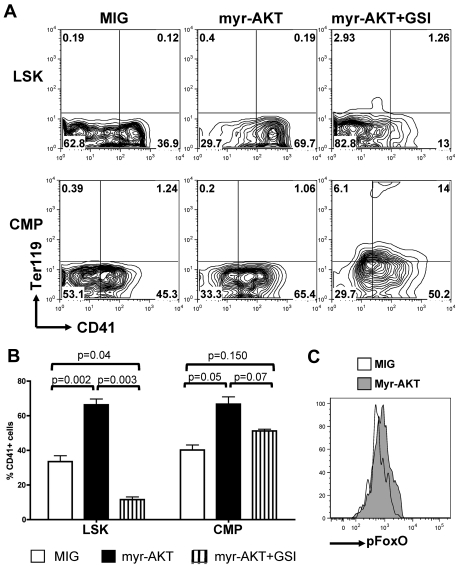
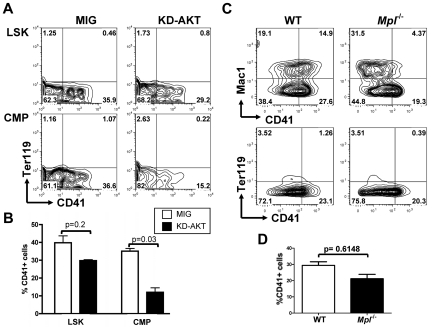
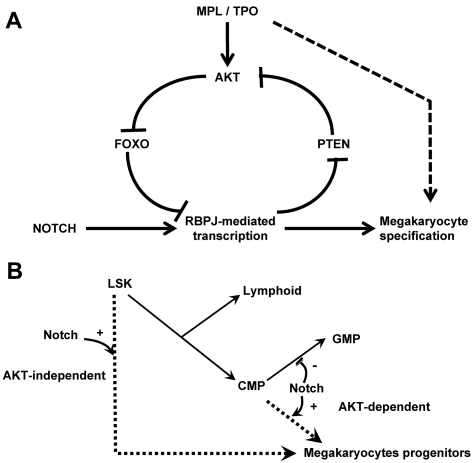
The NOTCH signaling pathway is implicated in a broad range of developmental processes, including cell fate decisions. However, the molecular basis for its role at the different steps of stem cell lineage commitment is unclear. We recently identified the NOTCH signaling pathway as a positive regulator of megakaryocyte lineage specification during hematopoiesis, but the developmental pathways that allow hematopoietic stem cell differentiation into the erythro-megakaryocytic lineages remain controversial. Here, we investigated the role of downstream mediators of NOTCH during megakaryopoiesis and report crosstalk between the NOTCH and PI3K/AKT pathways. We demonstrate the inhibitory role of phosphatase with tensin homolog and Forkhead Box class O factors on megakaryopoiesis in vivo. Finally, our data annotate developmental mechanisms in the hematopoietic system that enable a decision to be made either at the hematopoietic stem cell or the committed progenitor level to commit to the megakaryocyte lineage, supporting the existence of 2 distinct developmental pathways.
Figures
References
-
- Luc S, Buza-Vidas N, Jacobsen SE. Delineating the cellular pathways of hematopoietic lineage commitment. Semin Immunol. 2008;20(4):213–220. - PubMed
-
- Luc S, Anderson K, Kharazi S, et al. Down-regulation of Mpl marks the transition to lymphoid-primed multipotent progenitors with gradual loss of granulocyte-monocyte potential. Blood. 2008;111(7):3424–3434. - PubMed
-
- Pronk CJ, Rossi DJ, Mansson R, et al. Elucidation of the phenotypic, functional, and molecular topography of a myeloerythroid progenitor cell hierarchy. Cell Stem Cell. 2007;1(4):428–442. - PubMed
-
- Forsberg EC, Serwold T, Kogan S, Weissman IL, Passegue E. New evidence supporting megakaryocyte-erythrocyte potential of flk2/flt3+ multipotent hematopoietic progenitors. Cell. 2006;126(2):415–426. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
