

Interaction of the human prostacyclin receptor with the PDZ adapter protein PDZK1: role in endothelial cell migration and angiogenesis
- PMID: 21653824
- PMCID: PMC3145543
- DOI: 10.1091/mbc.E11-04-0374
Interaction of the human prostacyclin receptor with the PDZ adapter protein PDZK1: role in endothelial cell migration and angiogenesis
Abstract
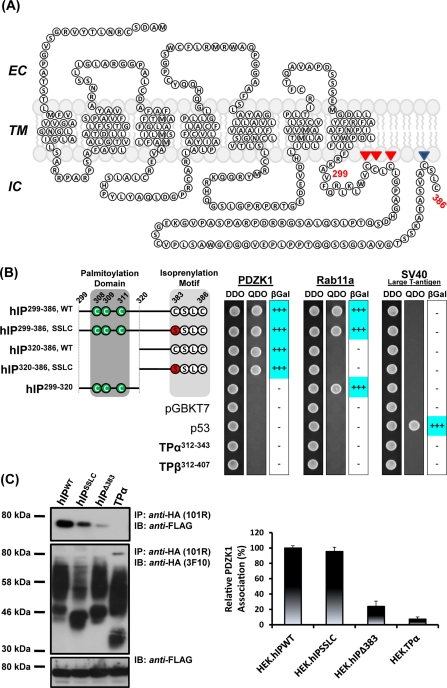
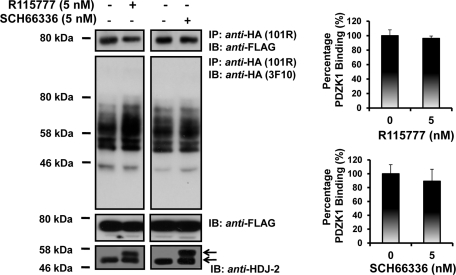
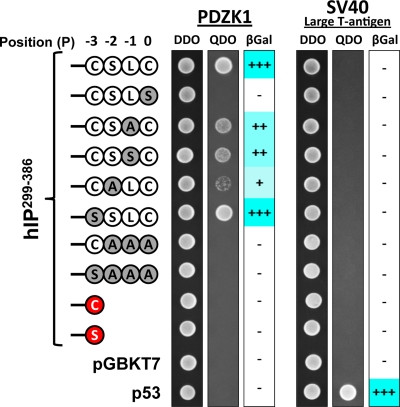
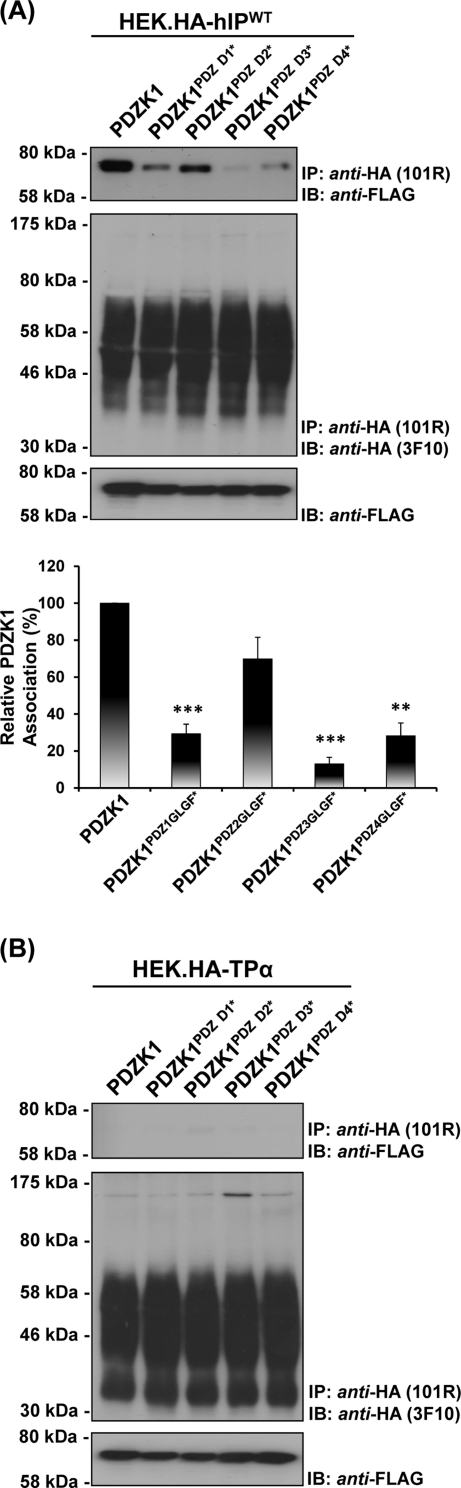
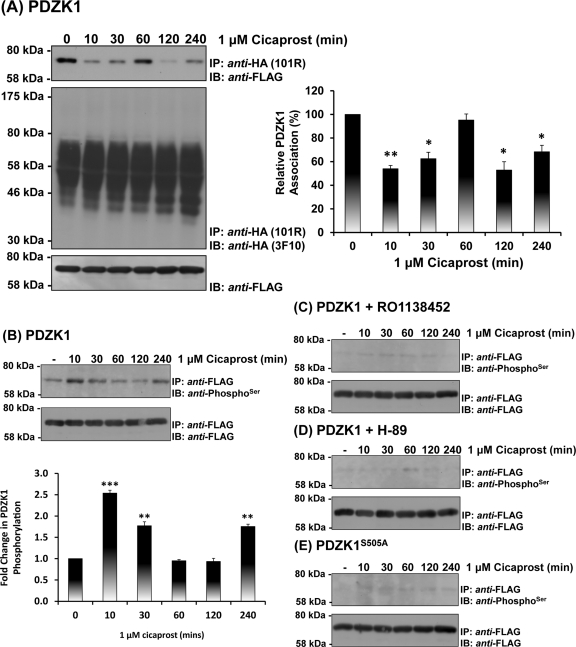
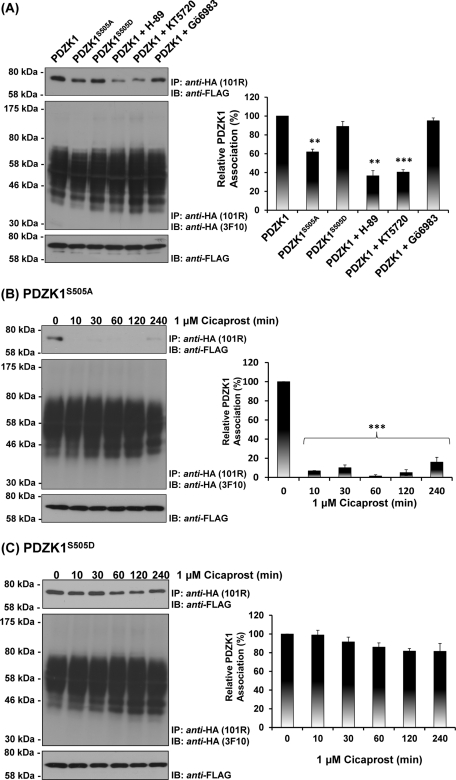
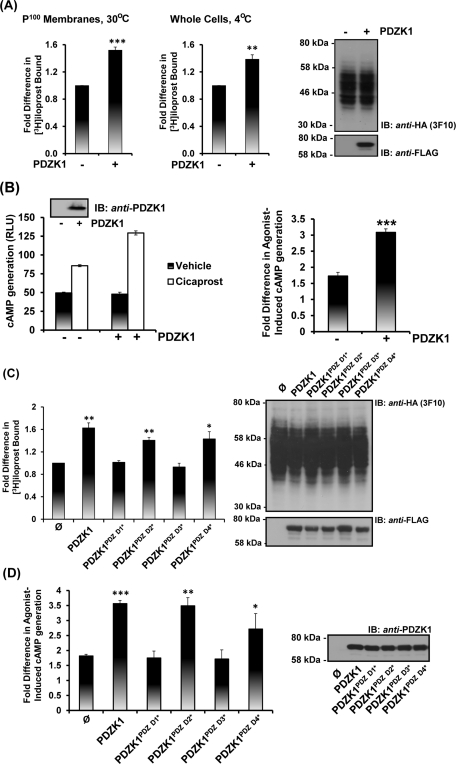
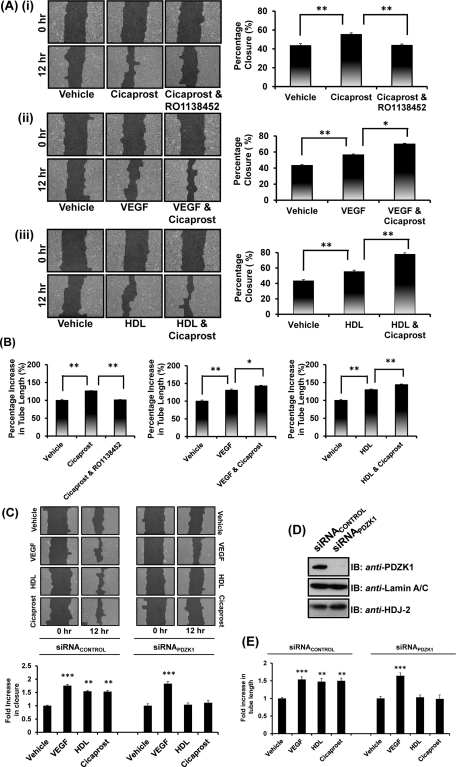
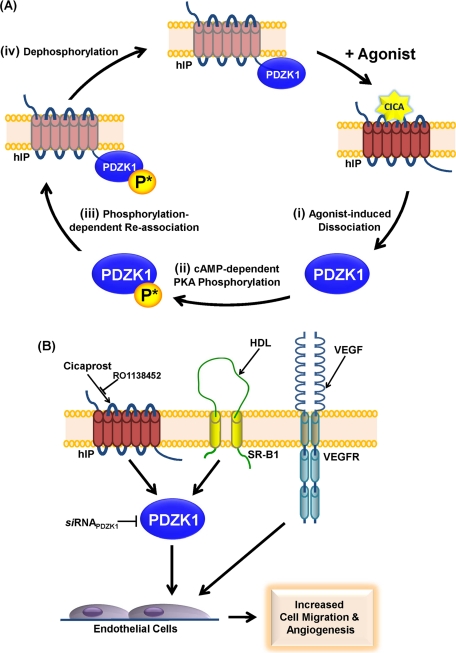
Prostacyclin is increasingly implicated in re-endothelialization and angiogenesis but through largely unknown mechanisms. Herein the high-density lipoprotein (HDL) scavenger receptor class B, type 1 (SR-B1) adapter protein PDZ domain-containing protein 1 (PDZK1) was identified as an interactant of the human prostacyclin receptor (hIP) involving a Class I PDZ ligand at its carboxyl terminus and PDZ domains 1, 3, and 4 of PDZK1. Although the interaction is constitutive, it may be dynamically regulated following cicaprost activation of the hIP through a mechanism involving cAMP-dependent protein kinase (PK)A-phosphorylation of PDZK1 at Ser-505. Although PDZK1 did not increase overall levels of the hIP, it increased its functional expression at the cell surface, enhancing ligand binding and cicaprost-induced cAMP generation. Consistent with its role in re-endothelialization and angiogenesis, cicaprost activation of the hIP increased endothelial cell migration and tube formation/in vitro angiogenesis, effects completely abrogated by the specific IP antagonist RO1138452. Furthermore, similar to HDL/SR-B1, small interfering RNA (siRNA)-targeted disruption of PDZK1 abolished cicaprost-mediated endothelial responses but did not affect VEGF responses. Considering the essential role played by prostacyclin throughout the cardiovascular system, identification of PDZK1 as a functional interactant of the hIP sheds significant mechanistic insights into the protective roles of these key players, and potentially HDL/SR-B1, within the vascular endothelium.
Figures
References
-
- Biscetti F, et al. Selective activation of peroxisome proliferator-activated receptor (PPAR)alpha and PPAR gamma induces neoangiogenesis through a vascular endothelial growth factor-dependent mechanism. Diabetes. 2008;57:1394–1404. - PubMed
-
- Biscetti F, et al. Peroxisome proliferator-activated receptor alpha is crucial for iloprost-induced in vivo angiogenesis and vascular endothelial growth factor upregulation. J Vasc Res. 2009;46:103–108. - PubMed
-
- Biscetti F, Pola R. Endothelial progenitor cells and angiogenesis join the PPARty. Circ Res. 2008;103:7–9. - PubMed
-
- Clark RD, Jahangir A, Severance D, Salazar R, Chang T, Chang D, Jett MF, Smith S, Bley K. Discovery and SAR development of 2-(phenylamino) imidazolines as prostacyclin receptor antagonists [corrected] Bioorg Med Chem Lett. 2004;14:1053–1056. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
