

A function for cyclin D1 in DNA repair uncovered by protein interactome analyses in human cancers
- PMID: 21654808
- PMCID: PMC3134411
- DOI: 10.1038/nature10155
A function for cyclin D1 in DNA repair uncovered by protein interactome analyses in human cancers
Abstract
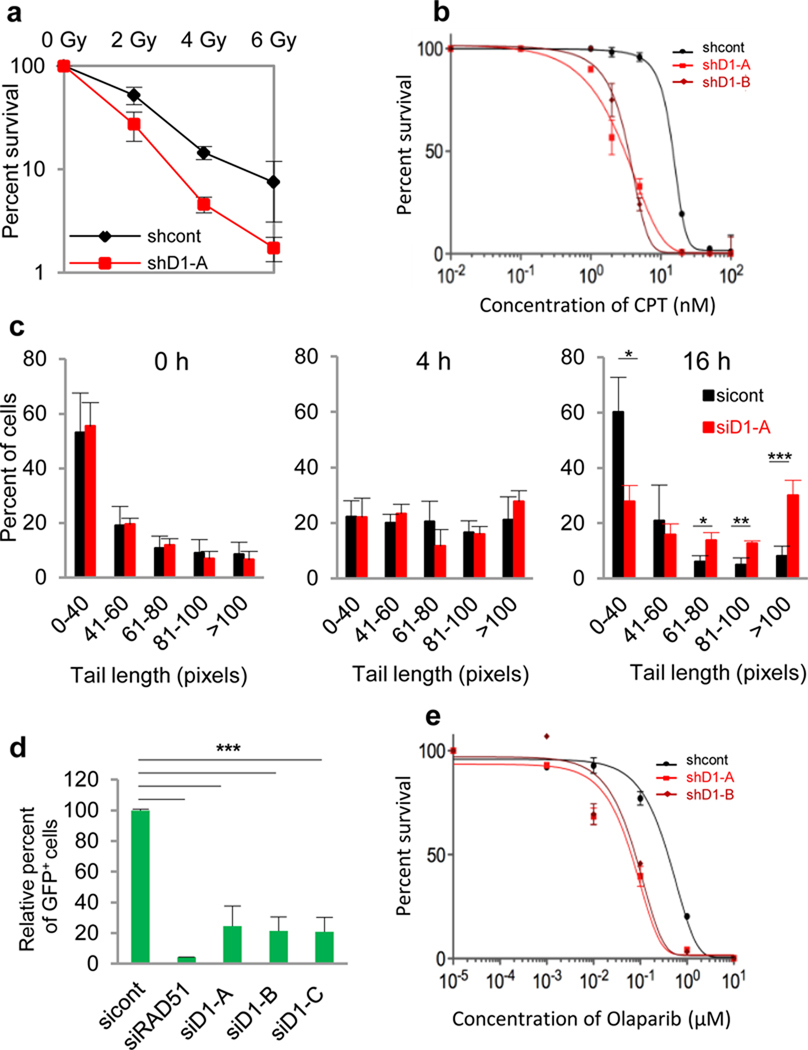
Cyclin D1 is a component of the core cell cycle machinery. Abnormally high levels of cyclin D1 are detected in many human cancer types. To elucidate the molecular functions of cyclin D1 in human cancers, we performed a proteomic screen for cyclin D1 protein partners in several types of human tumours. Analyses of cyclin D1 interactors revealed a network of DNA repair proteins, including RAD51, a recombinase that drives the homologous recombination process. We found that cyclin D1 directly binds RAD51, and that cyclin D1-RAD51 interaction is induced by radiation. Like RAD51, cyclin D1 is recruited to DNA damage sites in a BRCA2-dependent fashion. Reduction of cyclin D1 levels in human cancer cells impaired recruitment of RAD51 to damaged DNA, impeded the homologous recombination-mediated DNA repair, and increased sensitivity of cells to radiation in vitro and in vivo. This effect was seen in cancer cells lacking the retinoblastoma protein, which do not require D-cyclins for proliferation. These findings reveal an unexpected function of a core cell cycle protein in DNA repair and suggest that targeting cyclin D1 may be beneficial also in retinoblastoma-negative cancers which are currently thought to be unaffected by cyclin D1 inhibition.
Figures



Comment in
-
DNA repair: Cyclin D1 multitasks.Nature. 2011 Jun 8;474(7350):171-2. doi: 10.1038/474171a. Nature. 2011. PMID: 21654798 No abstract available.
References
-
- Sherr CJ, Roberts JM. Living with or without cyclins and cyclin-dependent kinases. Genes Dev. 2004;18:2699–2711. - PubMed
-
- Deshpande A, Sicinski P, Hinds PW. Cyclins and cdks in development and cancer: a perspective. Oncogene. 2005;24:2909–2915. - PubMed
-
- Baumann P, West SC. Role of the human RAD51 protein in homologous recombination and double-stranded-break repair. Trends Biochem Sci. 1998;23:247–251. - PubMed
-
- Bates S, et al. Absence of cyclin D/cdk complexes in cells lacking functional retinoblastoma protein. Oncogene. 1994;9:1633–1640. - PubMed
-
- Lukas J, et al. Retinoblastoma-protein-dependent cell-cycle inhibition by the tumour suppressor p16. Nature. 1995;375:503–506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
