

Voltage-gated channel mechanosensitivity: fact or friction?
- PMID: 21660289
- PMCID: PMC3107450
- DOI: 10.3389/fphys.2011.00025
Voltage-gated channel mechanosensitivity: fact or friction?
Abstract
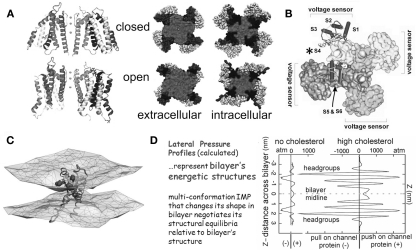
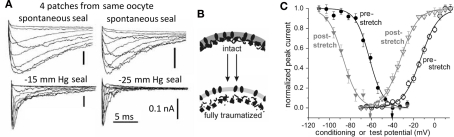
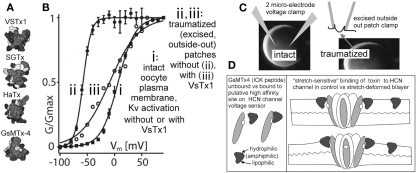
The heart is a continually active pulsatile fluid pump. It generates appropriate forces by precisely timed and spaced engagement of its contractile machinery. Largely, it makes its own control signals, the most crucial of which are precisely timed and spaced fluxes of ions across the sarcolemma, achieved by the timely opening and closing of diverse voltage-gated channels (VGC). VGCs have four voltage sensors around a central ion-selective pore that opens and closes under the influence of membrane voltage. Operation of any VGC is secondarily tuned by the mechanical state (i.e., structure) of the bilayer in which it is embedded. Rates of opening and closing, in other words, vary with bilayer structure. Thus, in the intensely mechanical environment of the myocardium and its vasculature, VGCs kinetics might be routinely modulated by reversible and irreversible nano-scale changes in bilayer structure. If subtle bilayer deformations are routine in the pumping heart, VGCs could be subtly transducing bilayer mechanical signals, thereby tuning cardiac rhythmicity, collectively contributing to mechano-electric feedback. Reversible bilayer deformations would be expected with changing shear flows and tissue distension, while irreversible bilayer restructuring occurs with ischemia, inflammation, membrane remodeling, etc. I suggest that tools now available could be deployed to help probe whether/how the inherent mechanosensitivity of VGCs - an attribute substantially reflecting the dependence of voltage sensor stability on bilayer structure - contributes to cardiac rhythmicity. Chief among these tools are voltage sensor toxins (whose inhibitory efficacy varies with the mechanical state of bilayer) and arrhythmia-inducing VGC mutants with distinctive mechano-phenotypes.
Keywords: LQT3; arrhythmias; bleb; ectopic excitation; mechano-electric feedback; pacemaker; sodium channel; stretch.
Figures



References
LinkOut - more resources
Full Text Sources