

Chromatin connections to pluripotency and cellular reprogramming
- PMID: 21663790
- PMCID: PMC4858411
- DOI: 10.1016/j.cell.2011.05.019
Chromatin connections to pluripotency and cellular reprogramming
Abstract
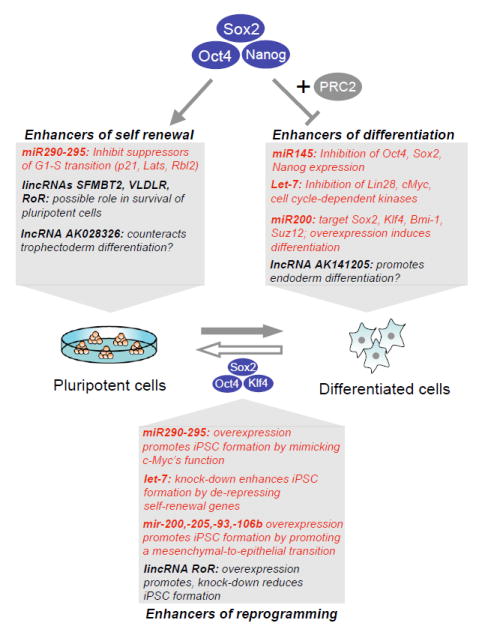
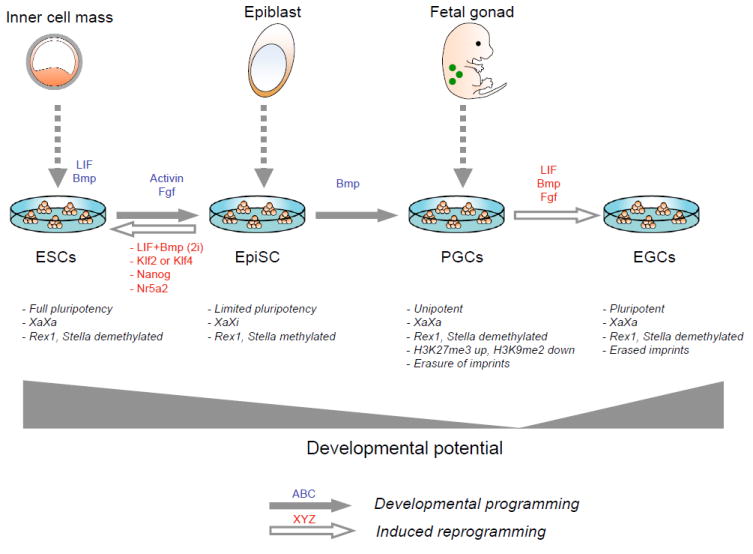
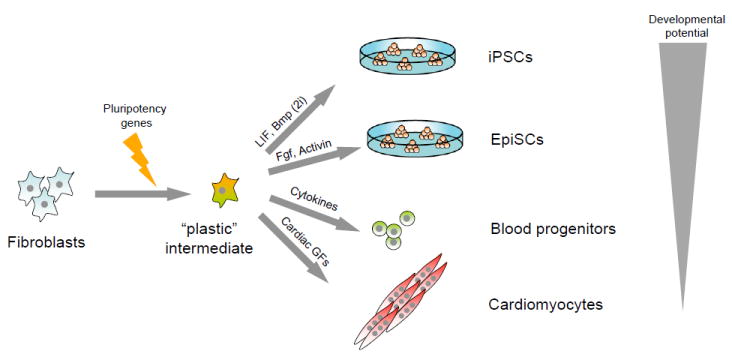
The pluripotent state of embryonic stem cells (ESCs) provides a unique perspective on regulatory programs that govern self-renewal and differentiation and somatic cell reprogramming. Here, we review the highly connected protein and transcriptional networks that maintain pluripotency and how they are intertwined with factors that affect chromatin structure and function. The complex interrelationships between pluripotency and chromatin factors are illustrated by X chromosome inactivation, regulatory control by noncoding RNAs, and environmental influences on cell states. Manipulation of cell state through the process of transdifferentiation suggests that environmental cues may direct transcriptional programs as cells enter a transiently "plastic" state during reprogramming.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
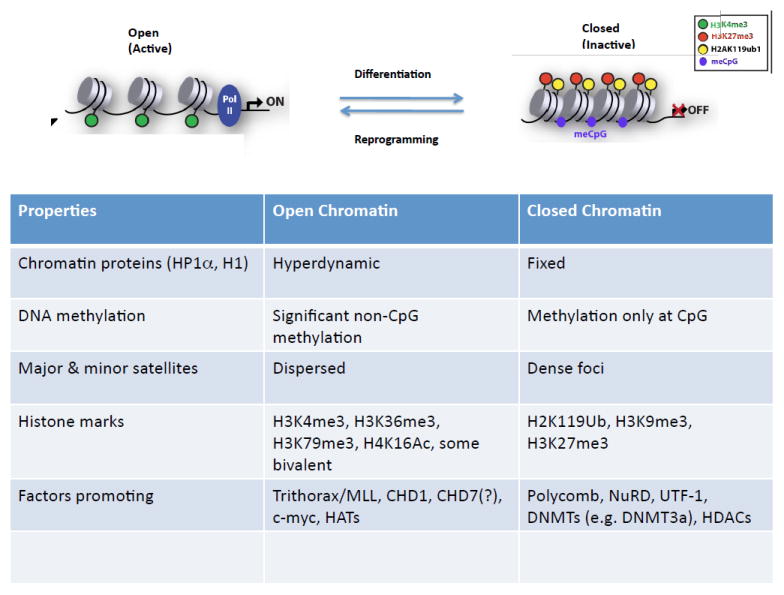
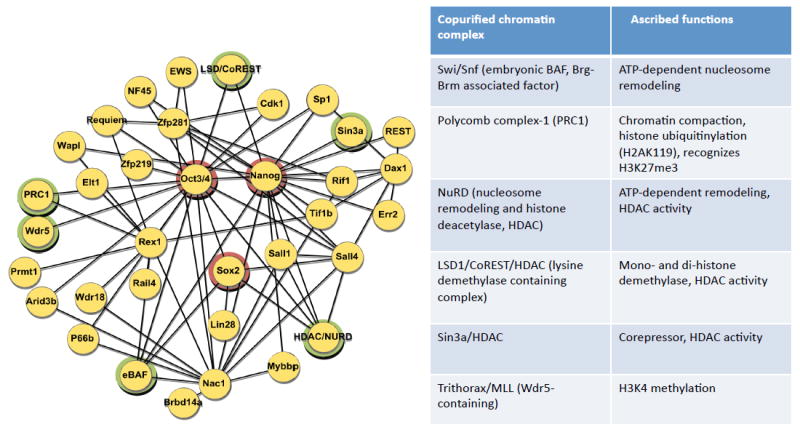
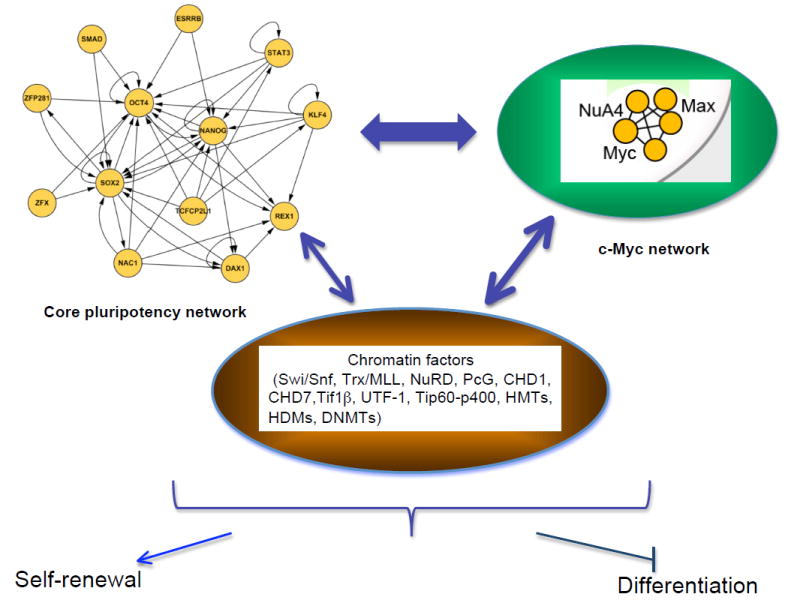
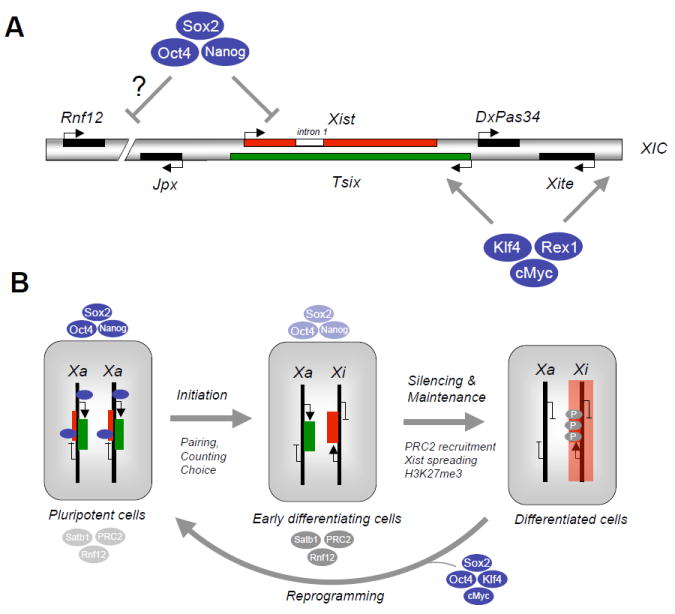
References
-
- Ancelin K, Lange UC, Hajkova P, Schneider R, Bannister AJ, Kouzarides T, Surani MA. Blimp1 associates with Prmt5 and directs histone arginine methylation in mouse germ cells. Nat Cell Biol. 2006;8:623–630. - PubMed
-
- Bantignies F, Roure V, Comet I, Leblanc B, Schuettengruber B, Bonnet J, Tixler V, Mas A, Cavalli G. Polycomb-dependent regulatoryu contacts between distant Hox loci in Drosophila. Cell. 2011;144:214–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
