

Vitamin D metabolism and function in the skin
- PMID: 21664236
- PMCID: PMC3188673
- DOI: 10.1016/j.mce.2011.05.017
Vitamin D metabolism and function in the skin
Abstract
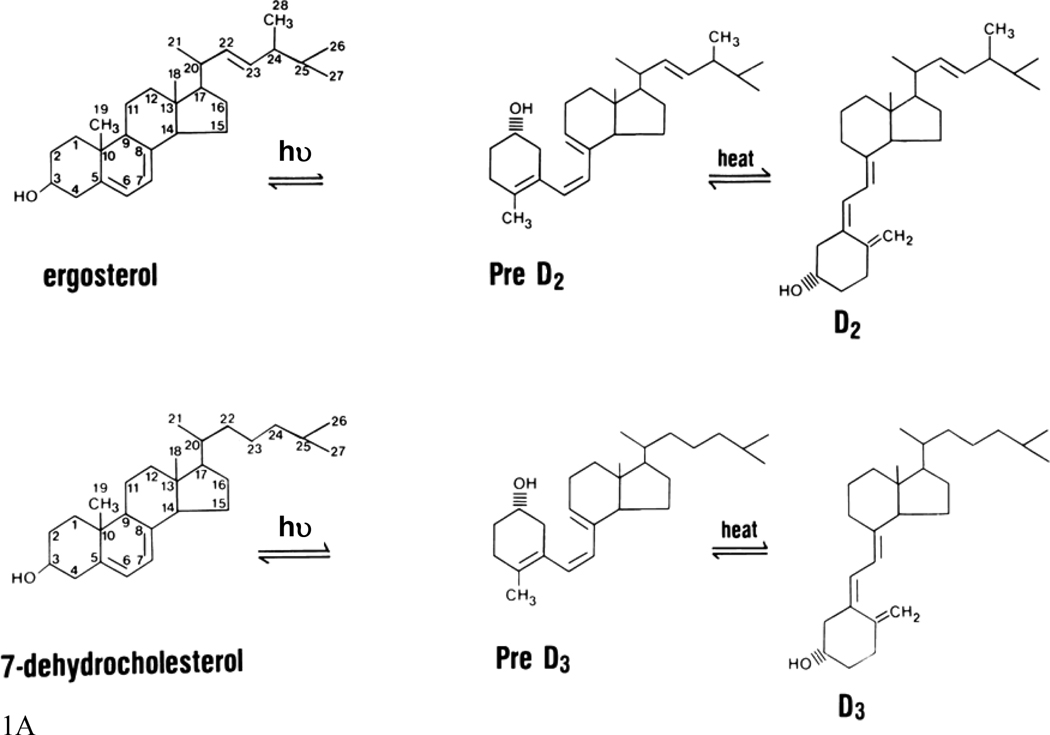
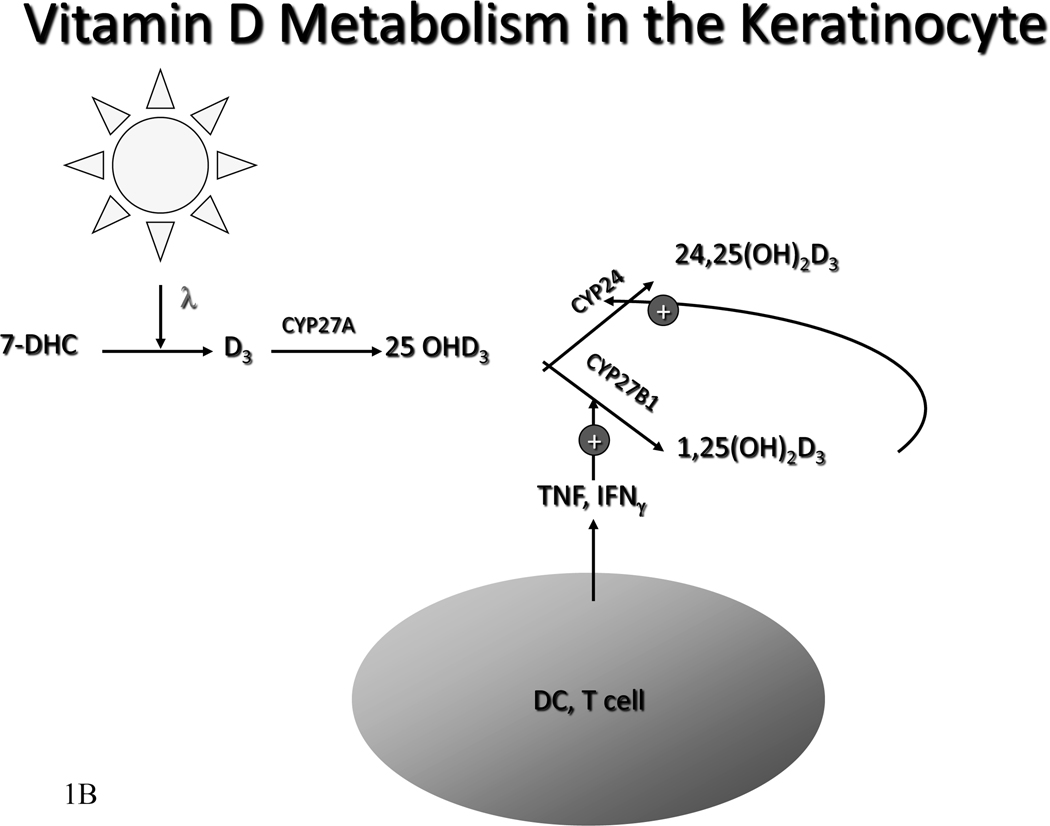
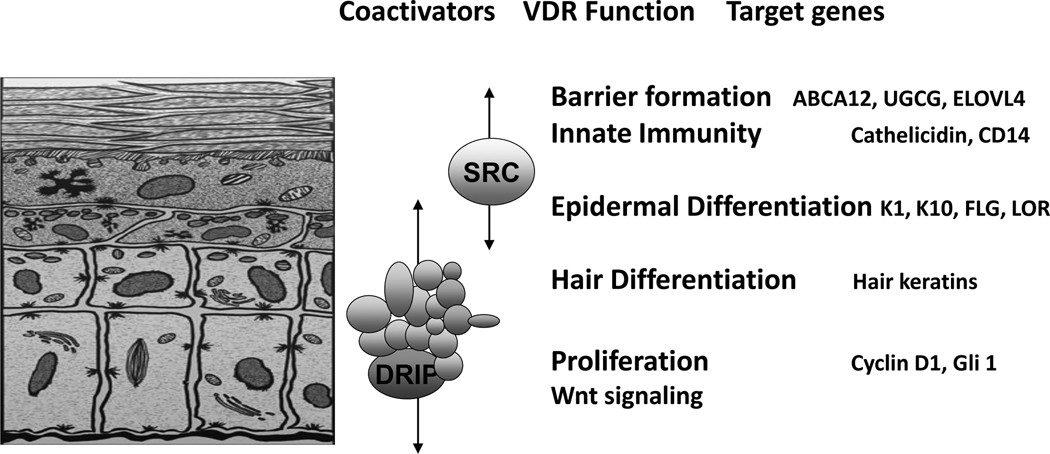
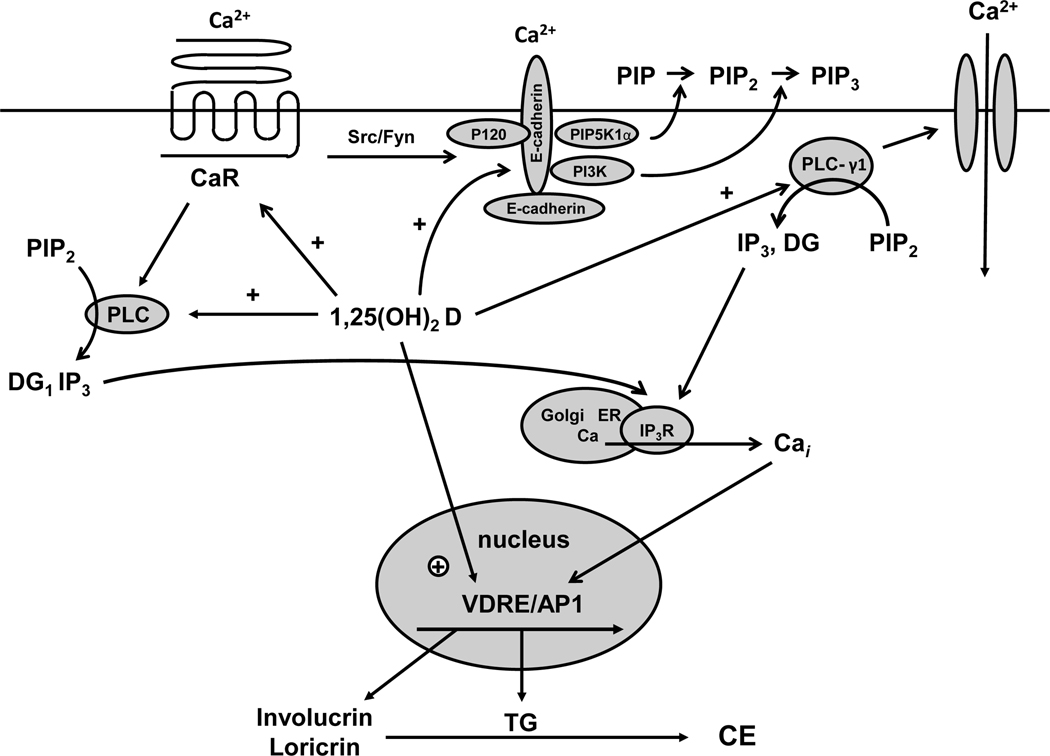
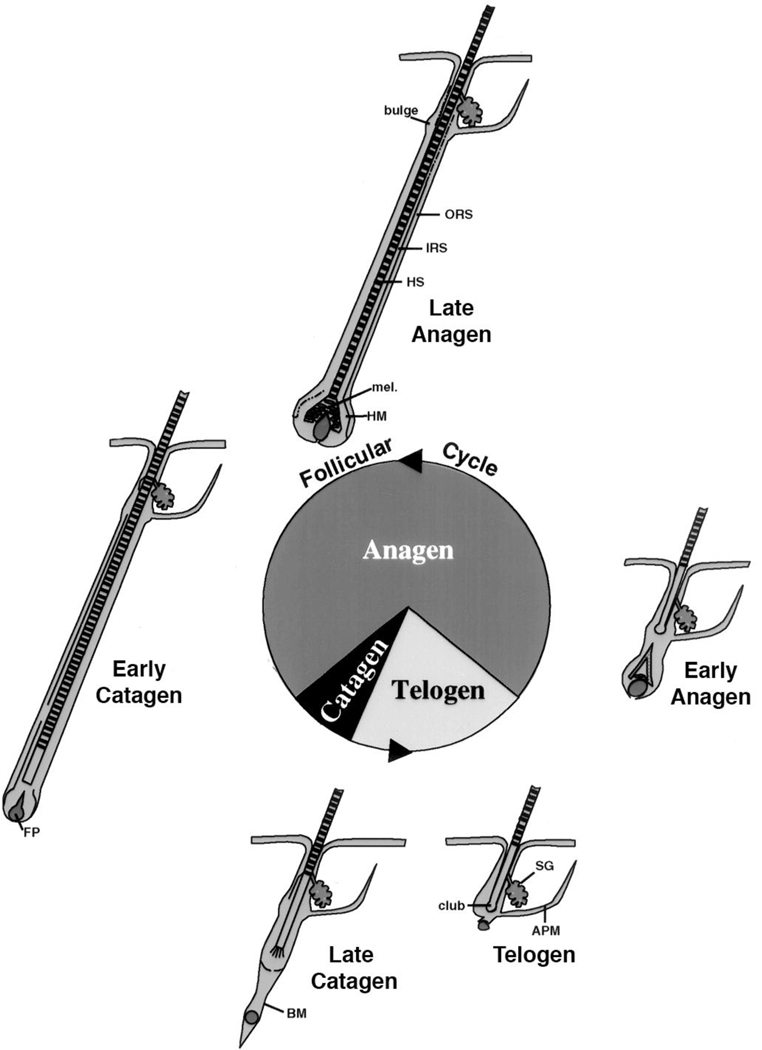
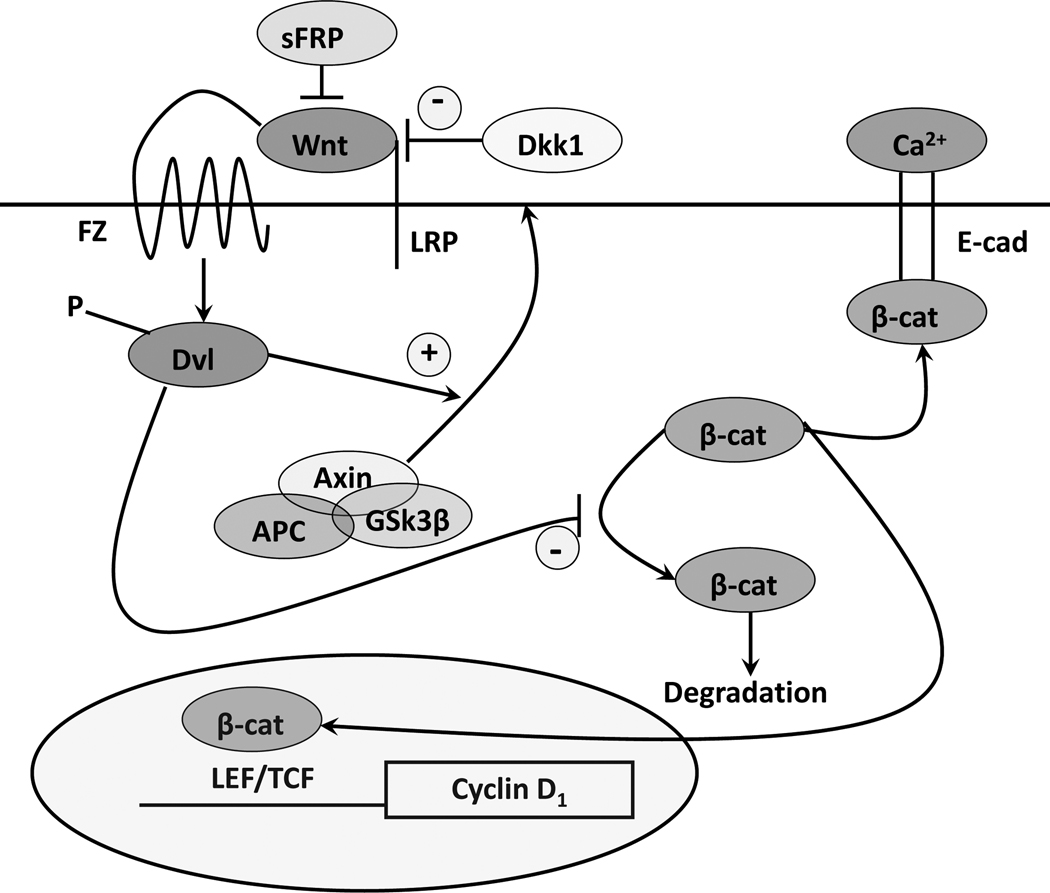
The keratinocytes of the skin are unique in being not only the primary source of vitamin D for the body, but in possessing the enzymatic machinery to metabolize vitamin D to its active metabolite 1,25(OH)(2)D. Furthermore, these cells also express the vitamin D receptor (VDR) that enables them to respond to the 1,25(OH)(2)D they produce. Numerous functions of the skin are regulated by 1,25(OH)(2)D and/or its receptor. These include inhibition of proliferation, stimulation of differentiation including formation of the permeability barrier, promotion of innate immunity, and promotion of the hair follicle cycle. Regulation of these actions is exerted by a number of different coregulators including the coactivators DRIP and SRC, the cosuppressor hairless (Hr), and β-catenin. This review will examine the regulation of vitamin D production and metabolism in the skin, and explore the various functions regulated by 1,25(OH)(2)D and its receptor.
Copyright © 2011 Elsevier Ireland Ltd. All rights reserved.
Figures
References
-
- Acevedo ML, Lee KC, Stender JD, et al. Selective recognition of distinct classes of coactivators by a ligand-inducible activation domain. Mol Cell. 2004;13:725–738. - PubMed
-
- Ahmad W, Faiyaz ul Haque M, Brancolini V, et al. Alopecia universalis associated with a mutation in the human hairless gene. Science. 1998;279:720–724. - PubMed
-
- Bienz M. beta-Catenin: a pivot between cell adhesion and Wnt signalling. Curr Biol. 2005;15:R64–R67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
