

Long-distance kissing loop interactions between a 3' proximal Y-shaped structure and apical loops of 5' hairpins enhance translation of Saguaro cactus virus
- PMID: 21664637
- PMCID: PMC3152624
- DOI: 10.1016/j.virol.2011.05.007
Long-distance kissing loop interactions between a 3' proximal Y-shaped structure and apical loops of 5' hairpins enhance translation of Saguaro cactus virus
Abstract
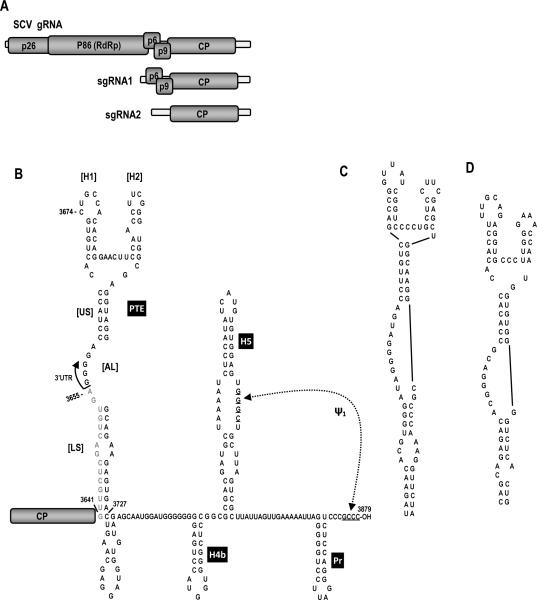
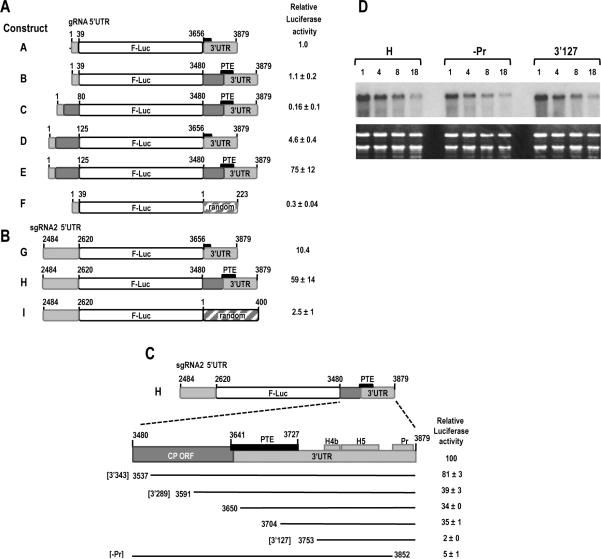
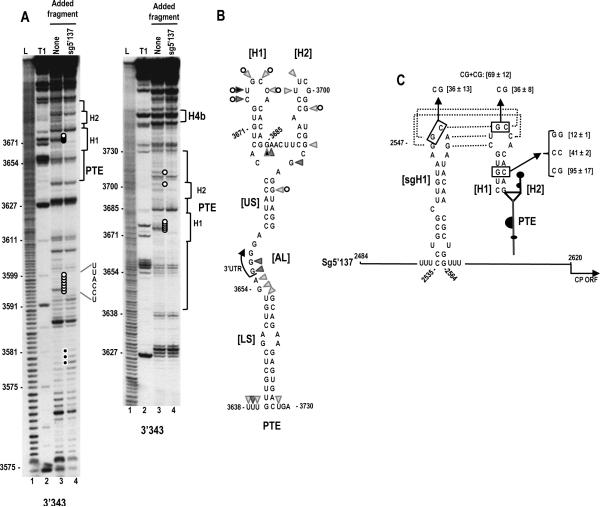
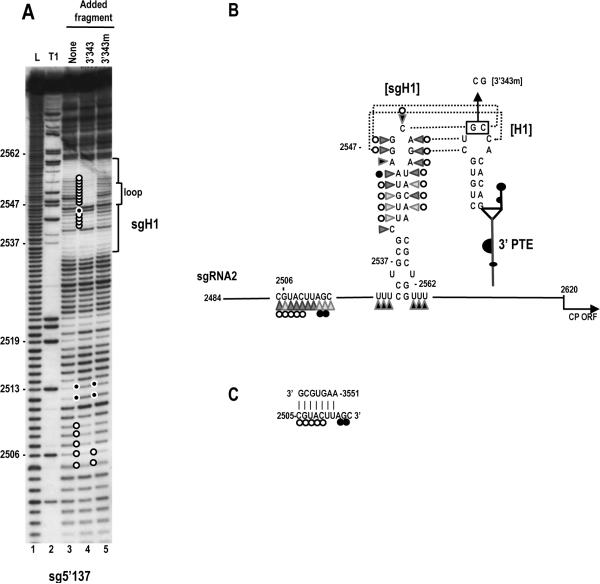
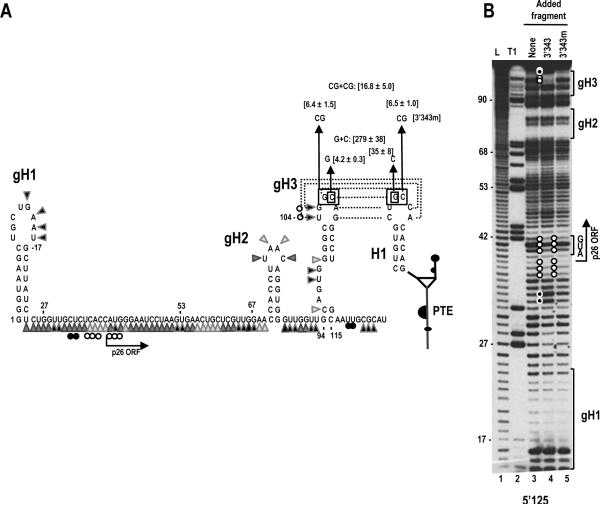
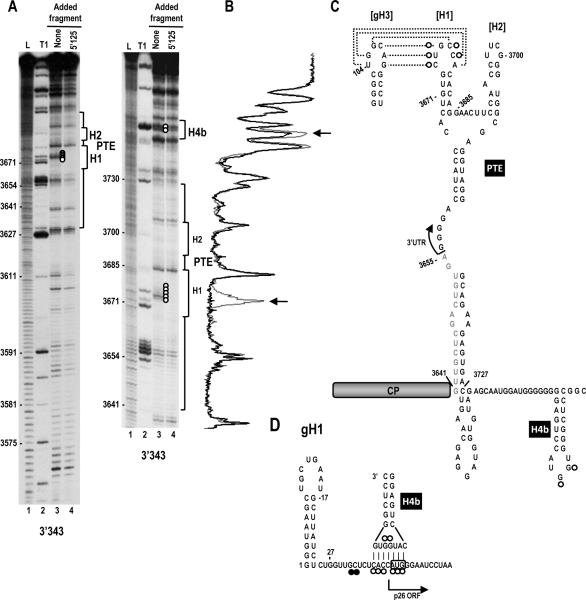
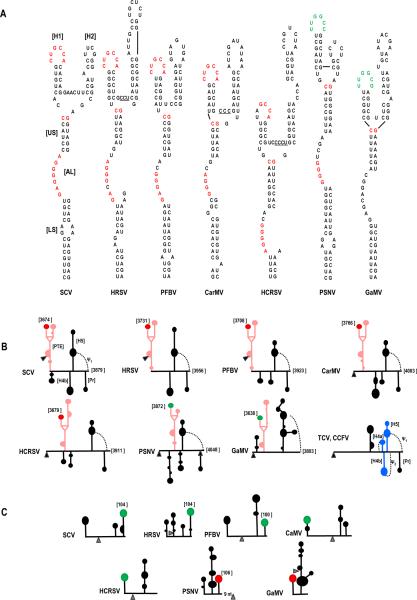
Circularization of cellular mRNAs is a key event prior to translation initiation. We report that efficient translation of Saguaro cactus virus (SCV) requires a 3' translational enhancer (PTE) located partially in coding sequences. Unlike a similar PTE reported in the 3' UTR of Pea enation mosaic virus that does not engage in an RNA:RNA interaction (Wang Z. et al., J. Biol. Chem. 284, 14189-14202, 2009), the SCV PTE participates in long distance RNA:RNA interactions with hairpins located in the p26 ORF and in the 5' UTR of one subgenomic RNA. At least two additional RNA:RNA interactions are also present, one of which involves the p26 initiation codon. Similar PTE can be found in six additional carmoviruses that can putatively form long-distance interactions with 5' hairpins located in comparable positions.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Batten JS, Desvoyes B, Yamamura Y, Scholthof KBG. A translational enhancer element on the 3'-proximal end of the Panicum mosaic virus genome. FEBS Let. 2006;580:2591–2597. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
