

An ALS-associated mutation affecting TDP-43 enhances protein aggregation, fibril formation and neurotoxicity
- PMID: 21666678
- PMCID: PMC3357956
- DOI: 10.1038/nsmb.2053
An ALS-associated mutation affecting TDP-43 enhances protein aggregation, fibril formation and neurotoxicity
Abstract
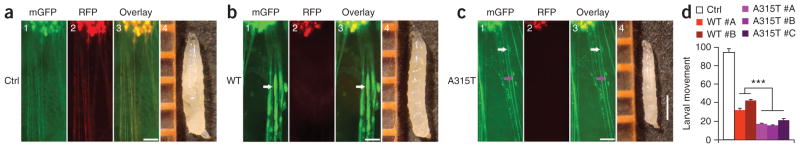
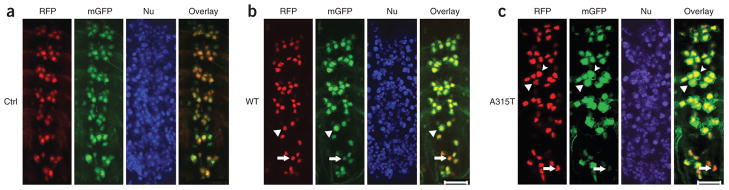
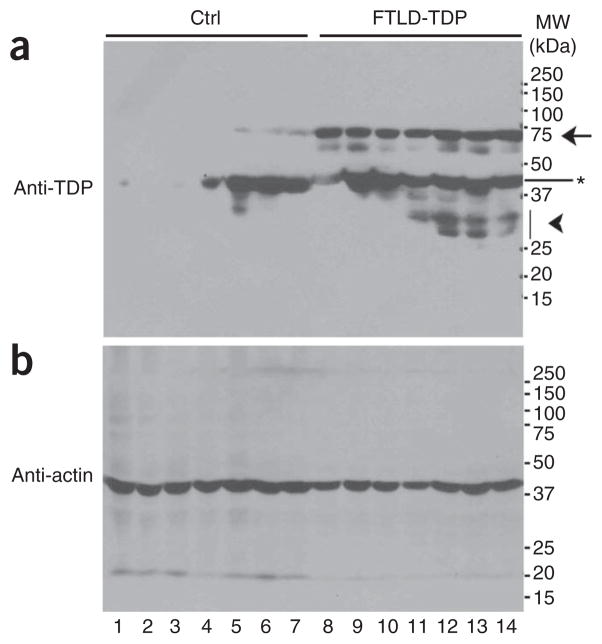
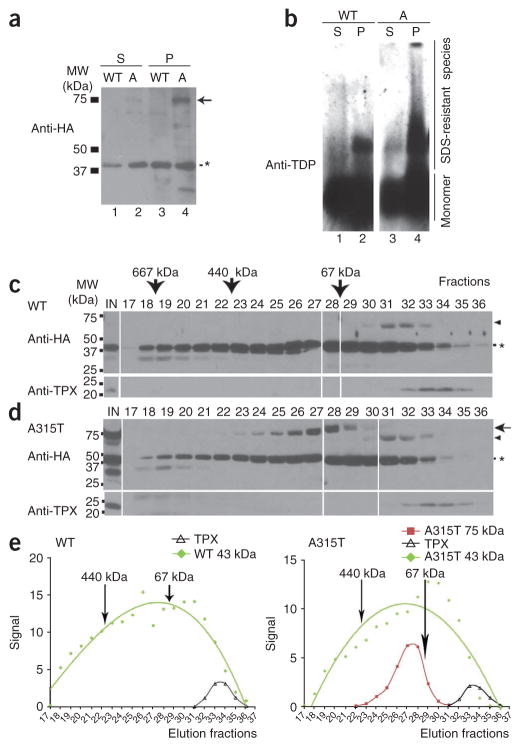
Mutations in TARDBP, encoding TAR DNA-binding protein-43 (TDP-43), are associated with TDP-43 proteinopathies, including amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration (FTLD). We compared wild-type TDP-43 and an ALS-associated mutant TDP-43 in vitro and in vivo. The A315T mutant enhances neurotoxicity and the formation of aberrant TDP-43 species, including protease-resistant fragments. The C terminus of TDP-43 shows sequence similarity to prion proteins. Synthetic peptides flanking residue 315 form amyloid fibrils in vitro and cause neuronal death in primary cultures. These data provide evidence for biochemical similarities between TDP-43 and prion proteins, raising the possibility that TDP-43 derivatives may cause spreading of the disease phenotype among neighboring neurons. Our work also suggests that decreasing the abundance of neurotoxic TDP-43 species, enhancing degradation or clearance of such TDP-43 derivatives and blocking the spread of the disease phenotype may have therapeutic potential for TDP-43 proteinopathies.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
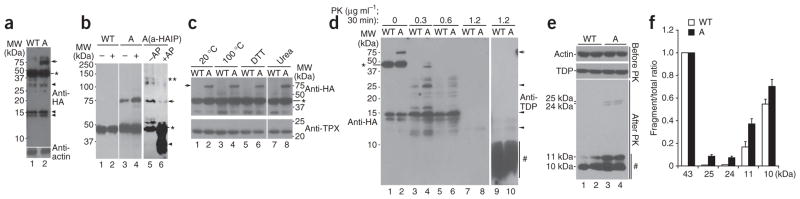
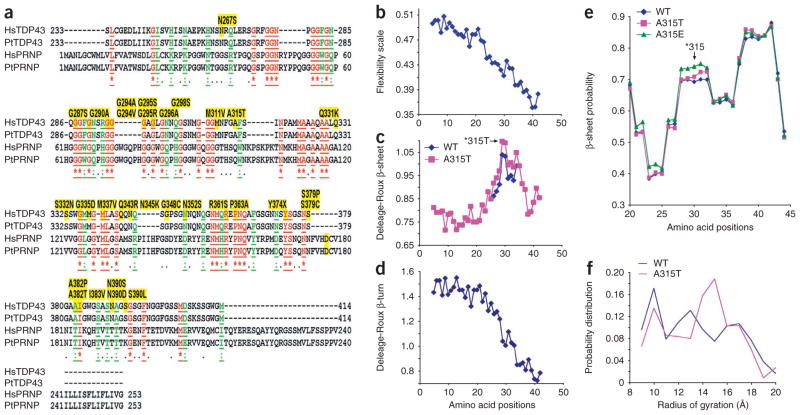
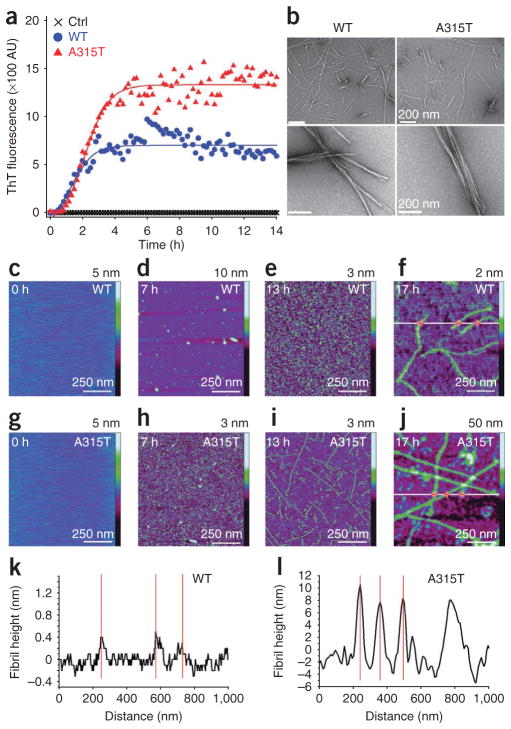
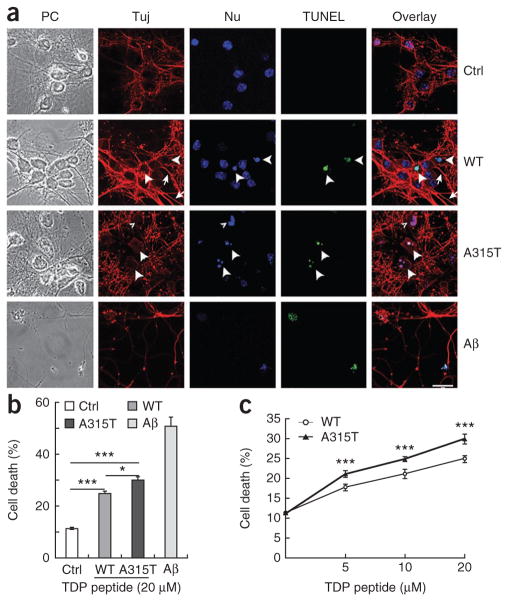
References
-
- Arai T, et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem Biophys Res Commun. 2006;351:602–611. - PubMed
-
- Neumann M, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–133. - PubMed
-
- Kwiatkowski TJ, Jr, et al. Mutations in the FUS/TLS gene on chromosome 16 cause familial amyotrophic lateral sclerosis. Science. 2009;323:1205–1208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
