

Superior antimalarial immunity after vaccination with late liver stage-arresting genetically attenuated parasites
- PMID: 21669394
- PMCID: PMC3117254
- DOI: 10.1016/j.chom.2011.05.008
Superior antimalarial immunity after vaccination with late liver stage-arresting genetically attenuated parasites
Abstract
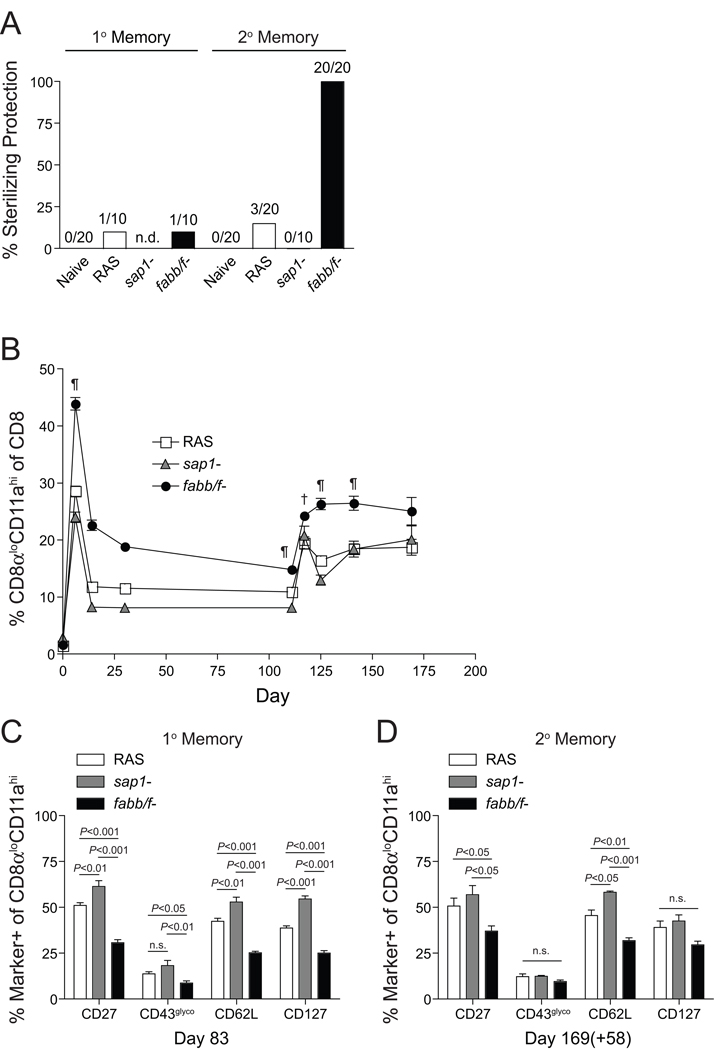
While subunit vaccines have shown partial efficacy in clinical trials, radiation-attenuated sporozoites (RAS) remain the "gold standard" for sterilizing protection against Plasmodium infection in human vaccinees. The variability in immunogenicity and replication introduced by the extensive, random DNA damage necessary to generate RAS could be overcome by genetically attenuated parasites (GAP) designed via gene deletion to arrest at defined points during liver-stage development. Here, we demonstrate the principle that late liver stage-arresting GAP induce larger and broader CD8 T cell responses that provide superior protection in inbred and outbred mice compared to RAS or early-arresting GAP immunizations. Late liver stage-arresting GAP also engender high levels of cross-stage and cross-species protection and complete protection when administered by translationally relevant intradermal or subcutaneous routes. Collectively, our results underscore the potential utility of late liver stage-arresting GAP as broadly protective next-generation live-attenuated malaria vaccines and support their potential as a powerful model for identifying antigens to generate cross-stage protection.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures




References
-
- Aide P, Aponte JJ, Renom M, Nhampossa T, Sacarlal J, Mandomando I, Bassat Q, Manaca MN, Leach A, Lievens M, et al. Safety, immunogenicity and duration of protection of the RTS,S/AS02(D) malaria vaccine: one year follow-up of a randomized controlled phase I/IIb trial. PLoS One. 2010;5:e13838. - PMC - PubMed
-
- Belnoue E, Voza T, Costa FT, Gruner AC, Mauduit M, Rosa DS, Depinay N, Kayibanda M, Vigario AM, Mazier D, et al. Vaccination with live Plasmodium yoelii blood stage parasites under chloroquine cover induces cross-stage immunity against malaria liver stage. J Immunol. 2008;181:8552–8558. - PMC - PubMed
-
- Berenzon D, Schwenk RJ, Letellier L, Guebre-Xabier M, Williams J, Krzych U. Protracted protection to Plasmodium berghei malaria is linked to functionally and phenotypically heterogeneous liver memory CD8+ T cells. J Immunol. 2003;171:2024–2034. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
