

Characterization of receptors for murine pregnancy specific glycoproteins 17 and 23
- PMID: 21669460
- PMCID: PMC3142296
- DOI: 10.1016/j.placenta.2011.05.008
Characterization of receptors for murine pregnancy specific glycoproteins 17 and 23
Abstract
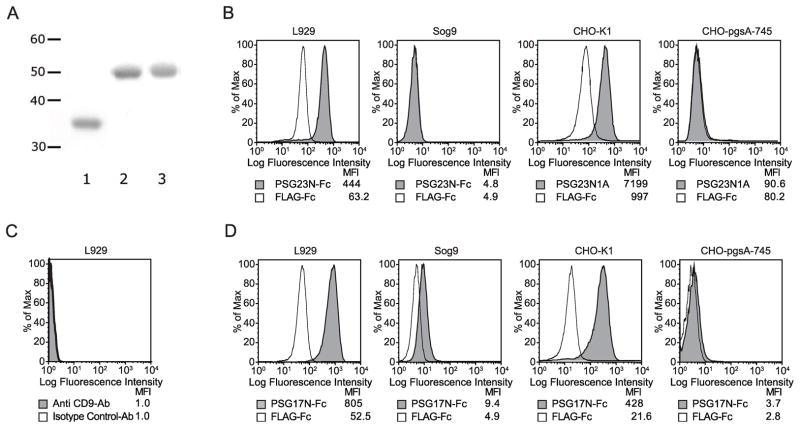
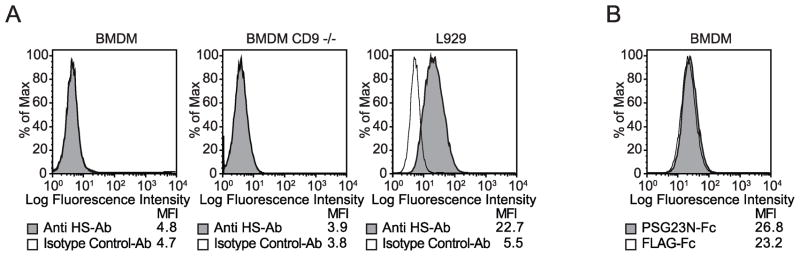
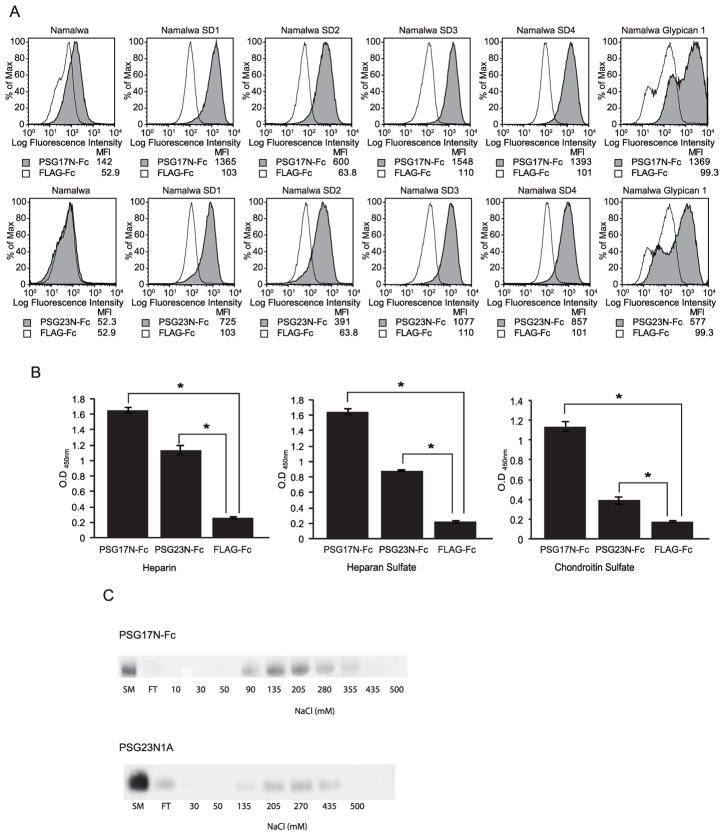
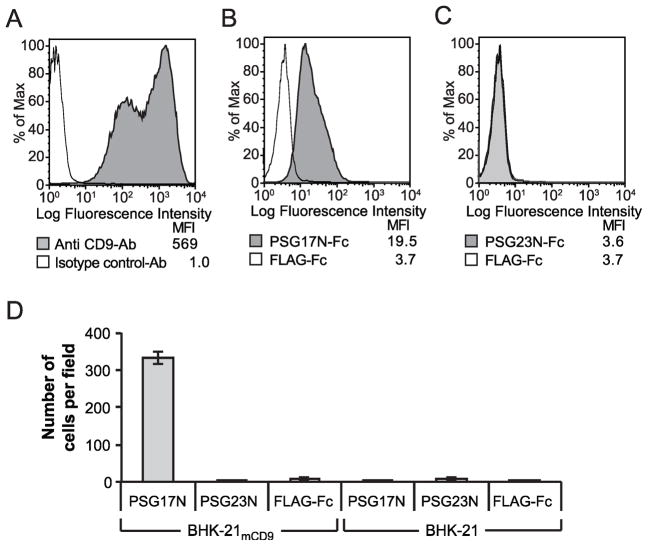
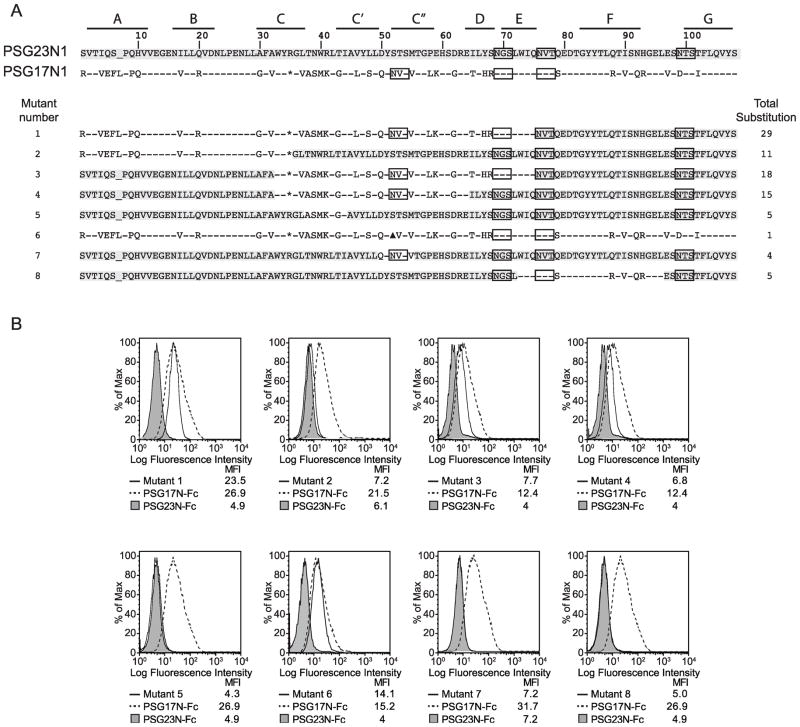
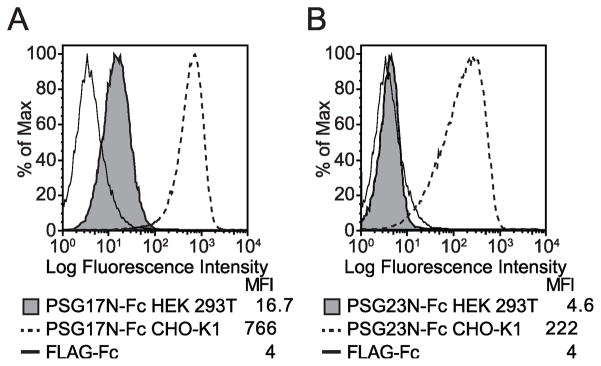
In primates and rodents, trophoblast cells synthesize and secrete into the maternal circulation a family of proteins known as pregnancy specific glycoproteins (PSG). The current study was undertaken to characterize the receptor for two members of the murine PSG family, PSG17 and PSG23. Binding of recombinant PSG17 and PSG23 to CHO-K1 and L929 cells and their derived mutants was performed to determine whether these proteins bound to cell surface proteoglycans. We also examined binding of these proteins to cells transfected with syndecans and glypican-1 by flow cytometry. The interaction with glycosaminoglycans was confirmed in solid phase assays. Our results show that PSG17 binds to CD9 and to cell surface proteoglycans while PSG23 binds only to the latter. We found that the amino acids involved in CD9 binding reside in the region of highest divergence between the N1-domains of murine PSGs. For both proteins, the N-terminal domain (designated as N1) is sufficient for binding to cells and the ability to bind cell surface proteoglycans is affected by the cell line employed to generate the recombinant proteins. We conclude that while substantially different at the amino acid level, some murine PSGs share with human PSG1 the ability to bind to cell surface proteoglycans and that at least one PSG binds to more than one type of molecule on the cell surface.
Published by Elsevier Ltd.
Figures
References
-
- Horne CH, Towler CM, Pugh-Humphreys RG, Thomson AW, Bohn H. Pregnancy specific beta1-glycoprotein--a product of the syncytiotrophoblast. Experientia. 1976;32(9):1197. - PubMed
-
- Kromer B, Finkenzeller D, Wessels J, Dveksler G, Thompson J, Zimmermann W. Coordinate expression of splice variants of the murine pregnancy-specific glycoprotein (PSG) gene family during placental development. Eur J Biochem. 1996;242(2):280–7. - PubMed
-
- Camolotto S, Racca A, Rena V, Nores R, Patrito LC, Genti-Raimondi S, et al. Expression and transcriptional regulation of individual pregnancy-specific glycoprotein genes in differentiating trophoblast cells. Placenta. 2010;31(4):312–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
