

Proteolytically cleaved MLL subunits are susceptible to distinct degradation pathways
- PMID: 21670200
- PMCID: PMC3113671
- DOI: 10.1242/jcs.080523
Proteolytically cleaved MLL subunits are susceptible to distinct degradation pathways
Abstract
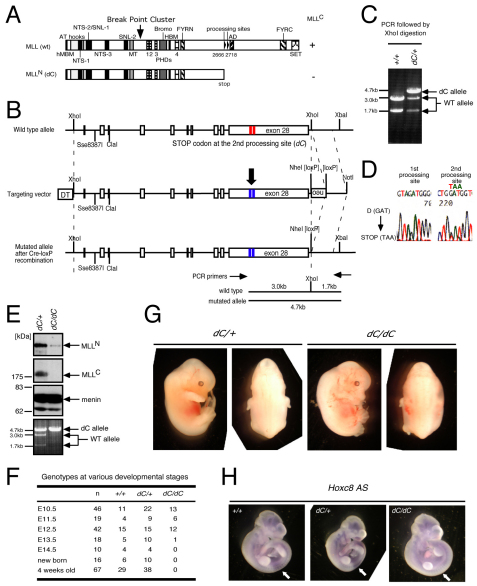
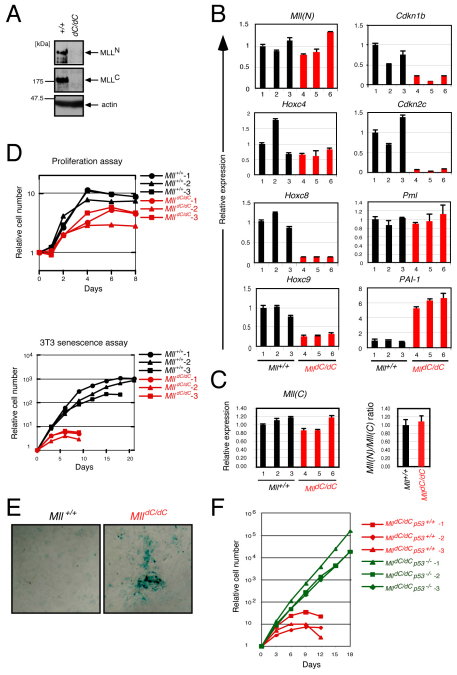
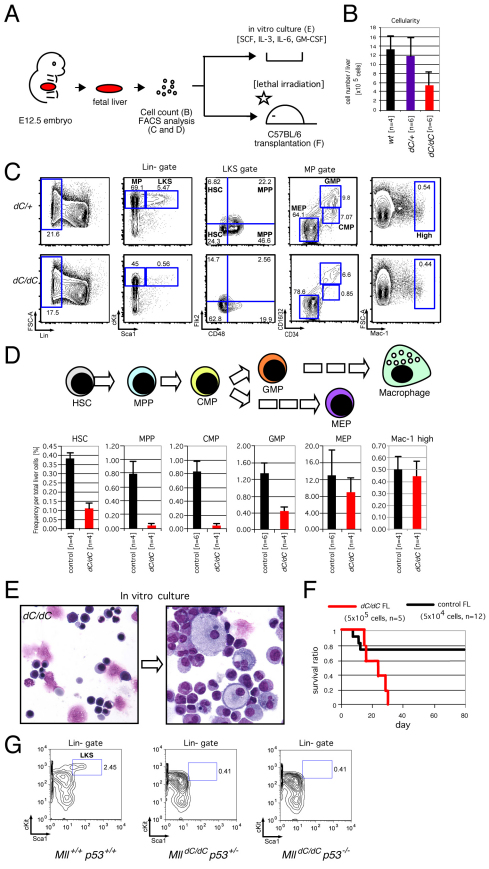
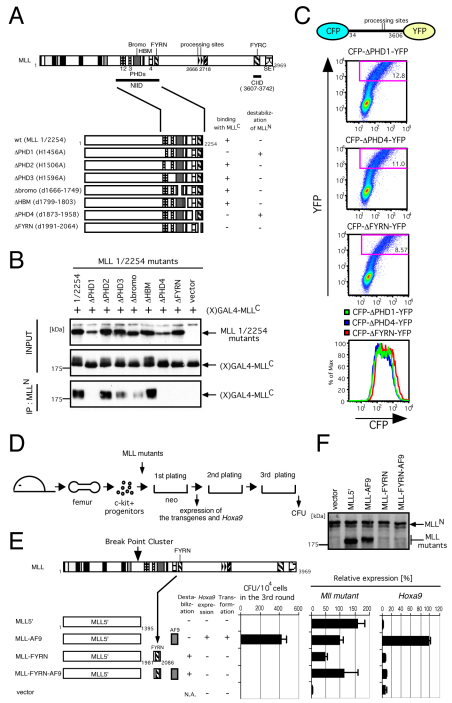
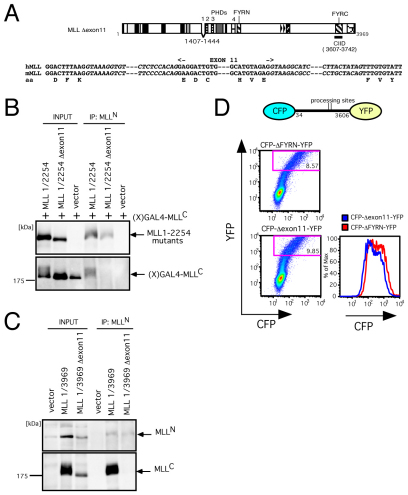
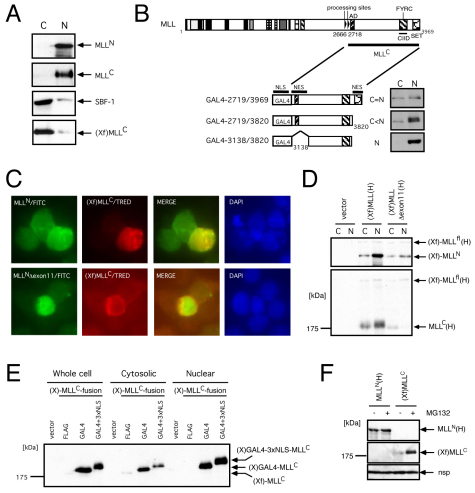
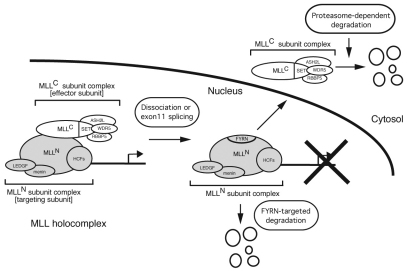
The mixed lineage leukemia (MLL) proto-oncogenic protein is a histone-lysine N-methyltransferase that is produced by proteolytic cleavage and self-association of the respective functionally distinct subunits (MLL(N) and MLL(C)) to form a holocomplex involved in epigenetic transcriptional regulation. On the basis of studies in Drosophila it has been suggested that the separated subunits might also have distinct functions. In this study, we used a genetically engineered mouse line that lacked MLL(C) to show that the MLL(N)-MLL(C) holocomplex is responsible for MLL functions in various developmental processes. The stability of MLL(N) is dependent on its intramolecular interaction with MLL(C), which is mediated through the first and fourth plant homeodomain (PHD) fingers (PHD1 and PHD4) and the phenylalanine/tyrosine-rich (FYRN) domain of MLL(N). Free MLL(N) is destroyed by a mechanism that targets the FYRN domain, whereas free MLL(C) is exported to the cytoplasm and degraded by the proteasome. PHD1 is encoded by an alternatively spliced exon that is occasionally deleted in T-cell leukemia, and its absence produces an MLL mutant protein that is deficient for holocomplex formation. Therefore, this should be a loss-of-function mutant allele, suggesting that the known tumor suppression role of MLL may also apply to the T-cell lineage. Our data demonstrate that the dissociated MLL subunits are subjected to distinct degradation pathways and thus not likely to have separate functions unless the degradation mechanisms are inhibited.
Figures
References
-
- Alvarez-Venegas R., Avramova Z. (2001). Two Arabidopsis homologs of the animal trithorax genes: a new structural domain is a signature feature of the trithorax gene family. Gene 271, 215-221 - PubMed
-
- Ayton P. M., Cleary M. L. (2001). Molecular mechanisms of leukemogenesis mediated by MLL fusion proteins. Oncogene 20, 5695-5707 - PubMed
-
- Bertolino P., Tong W. M., Galendo D., Wang Z. Q., Zhang C. X. (2003). Heterozygous Men1 mutant mice develop a range of endocrine tumors mimicking multiple endocrine neoplasia type 1. Mol. Endocrinol. 17, 1880-1892 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
