

BCL-2 modifying factor (BMF) is a central regulator of anoikis in human intestinal epithelial cells
- PMID: 21673109
- PMCID: PMC3143618
- DOI: 10.1074/jbc.M111.265322
BCL-2 modifying factor (BMF) is a central regulator of anoikis in human intestinal epithelial cells
Abstract
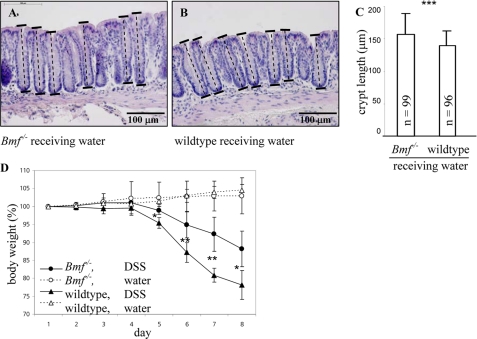
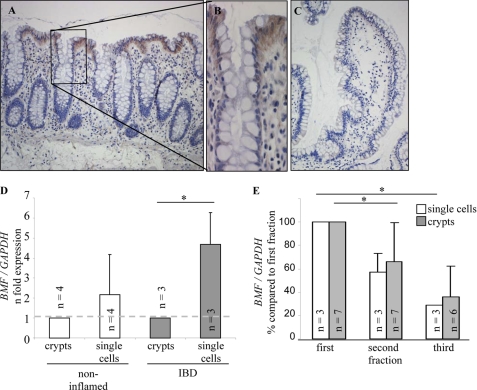
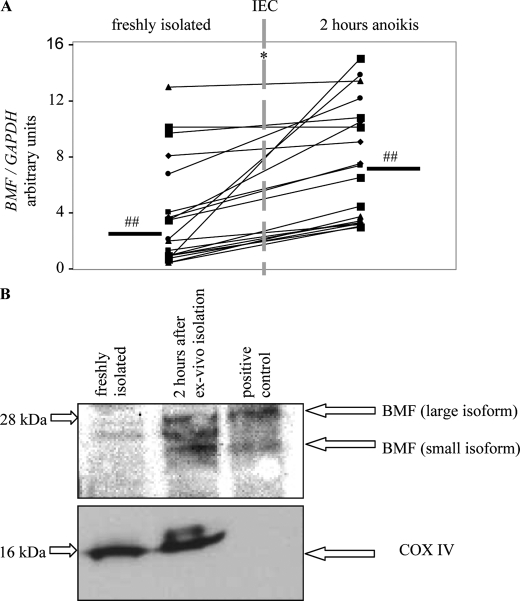
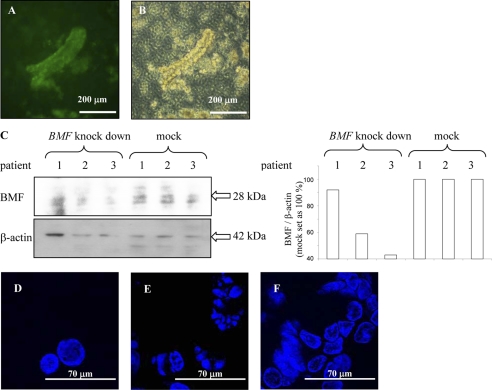
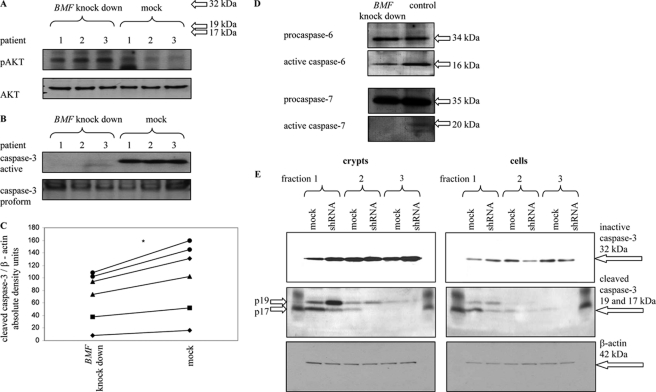
BCL-2 modifying factor (BMF) is a sentinel considered to register damage at the cytoskeleton and to convey a death signal to B-cell lymphoma 2. B-cell lymphoma 2 is neutralized by BMF and thereby facilitates cytochrome C release from mitochondria. We investigated the role of BMF for intestinal epithelial cell (IEC) homeostasis. Acute colitis was induced in Bmf-deficient mice (Bmf(-/-)) with dextran sulfate sodium. Colonic crypt length in Bmf(-/-) mice was significantly increased as compared with WT mice. Dextran sulfate sodium induced less signs of colitis in Bmf(-/-) mice, as weight loss was reduced compared with the WT. Primary human IEC exhibited increased BMF in the extrusion zone. Quantitative PCR showed a significant up-regulation of BMF expression after initiation of anoikis in primary human IEC. BMF was found on mitochondria during anoikis, as demonstrated by Western blot analysis. RNAi mediated knockdown of BMF reduced the number of apoptotic cells and led to reduced caspase 3 activity. A significant increase in phospho-AKT was determined after RNAi treatment. BMF knockdown supports survival of IEC. BMF is induced in human IEC by the loss of cell attachment and is likely to play an important role in the regulation of IEC survival.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
