

Cytotoxicity of farnesyltransferase inhibitors in lymphoid cells mediated by MAPK pathway inhibition and Bim up-regulation
- PMID: 21673341
- PMCID: PMC3208295
- DOI: 10.1182/blood-2011-02-334870
Cytotoxicity of farnesyltransferase inhibitors in lymphoid cells mediated by MAPK pathway inhibition and Bim up-regulation
Abstract
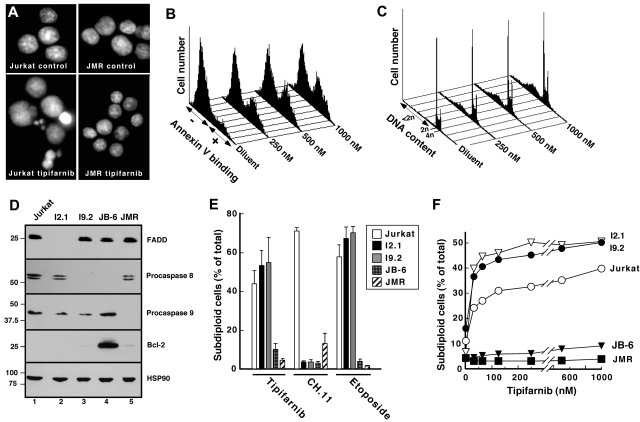
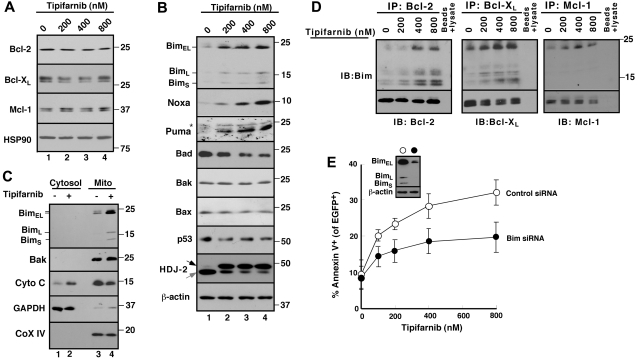
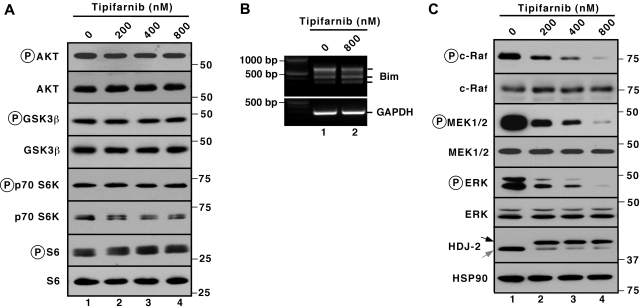
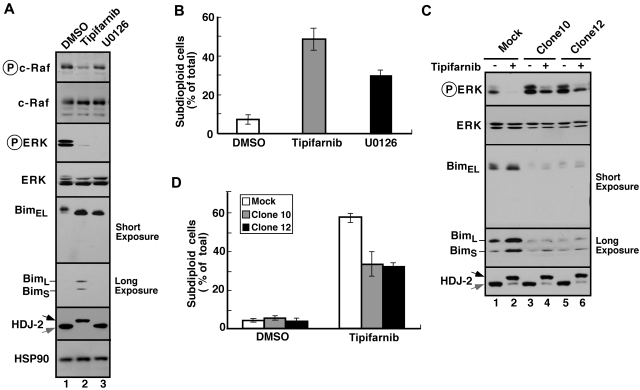
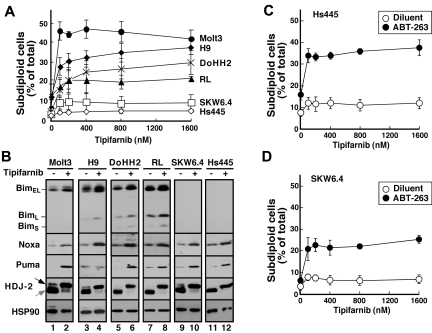
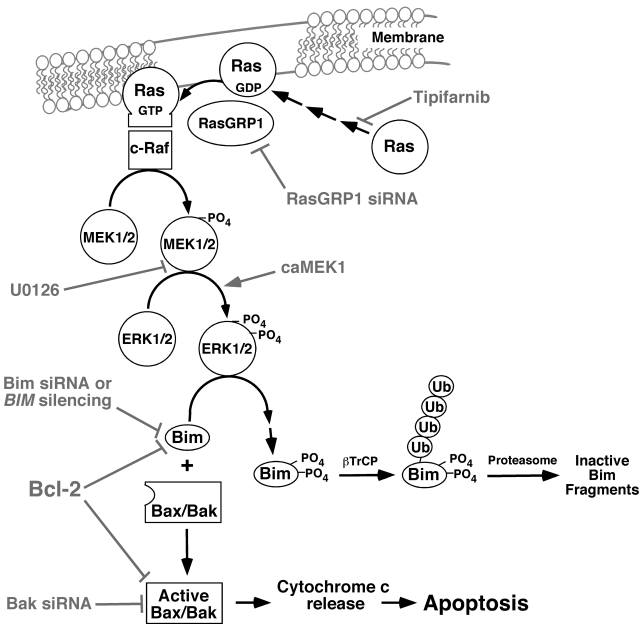
The mechanism of cytotoxicity of farnesyltransferase inhibitors is incompletely understood and seems to vary depending on the cell type. To identify potential determinants of sensitivity or resistance for study in the accompanying clinical trial (Witzig et al, page 4882), we examined the mechanism of cytotoxicity of tipifarnib in human lymphoid cell lines. Based on initial experiments showing that Jurkat variants lacking Fas-associated death domain or procaspase-8 undergo tipifarnib-induced apoptosis, whereas cells lacking caspase-9 or overexpressing Bcl-2 do not, we examined changes in Bcl-2 family members. Tipifarnib caused dose-dependent up-regulation of Bim in lymphoid cell lines (Jurkat, Molt3, H9, DoHH2, and RL) that undergo tipifarnib-induced apoptosis but not in lines (SKW6.4 and Hs445) that resist tipifarnib-induced apoptosis. Further analysis demonstrated that increased Bim levels reflect inhibition of signaling from c-Raf to MEK1/2 and ERK1/2. Additional experiments showed that down-regulation of the Ras guanine nucleotide exchange factor RasGRP1 diminished tipifarnib sensitivity, suggesting that H-Ras or N-Ras is a critical farnesylation target upstream of c-Raf in lymphoid cells. These results not only trace a pathway through c-Raf to Bim that contributes to tipifarnib cytotoxicity in human lymphoid cells but also identify potential determinants of sensitivity to this agent.
Figures

References
-
- Karp JE, Lancet JE. Development of farnesyltransferase inhibitors for clinical cancer therapy: focus on hematologic malignancies. Cancer Invest. 2007;25(6):484–494. - PubMed
-
- Harousseau JL. Farnesyltransferase inhibitors in hematologic malignancies. Blood Rev. 2007;21(4):173–182. - PubMed
-
- Stone RM. Targeted agents in AML: much more to do. Best Pract Res Clin Haematol. 2007;20(1):39–48. - PubMed
-
- Prendergast GC, Rane N. Farnesyltransferase inhibitors: mechanism and applications. Expert Opin Investig Drugs. 2001;10(12):2105–2116. - PubMed
-
- Sebti SM, Adjei AA. Farnesyltransferase inhibitors. Semin Oncol. 2004;31(1 suppl 1):28–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
