

β-Arrestin-mediated receptor trafficking and signal transduction
- PMID: 21680031
- PMCID: PMC3159699
- DOI: 10.1016/j.tips.2011.05.002
β-Arrestin-mediated receptor trafficking and signal transduction
Abstract
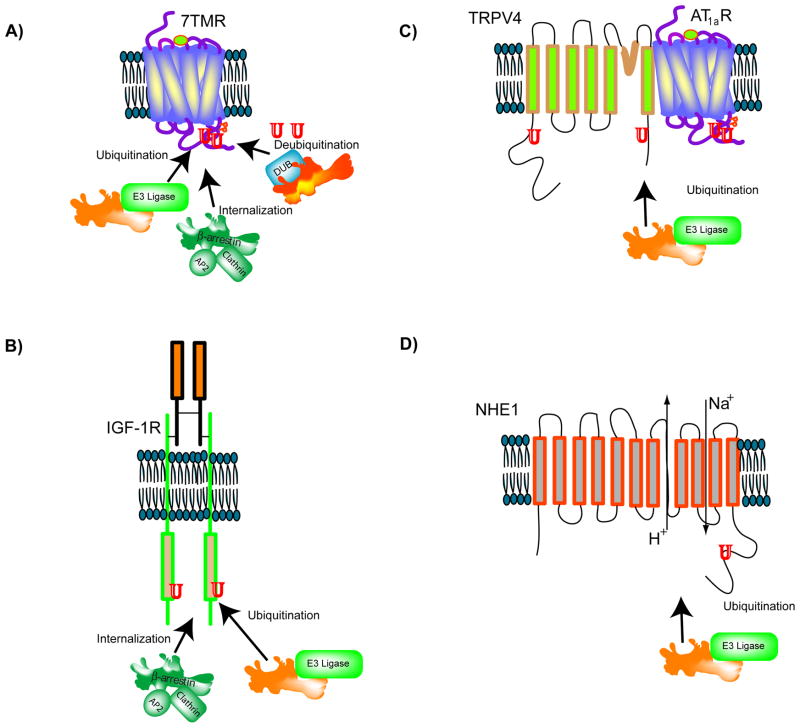
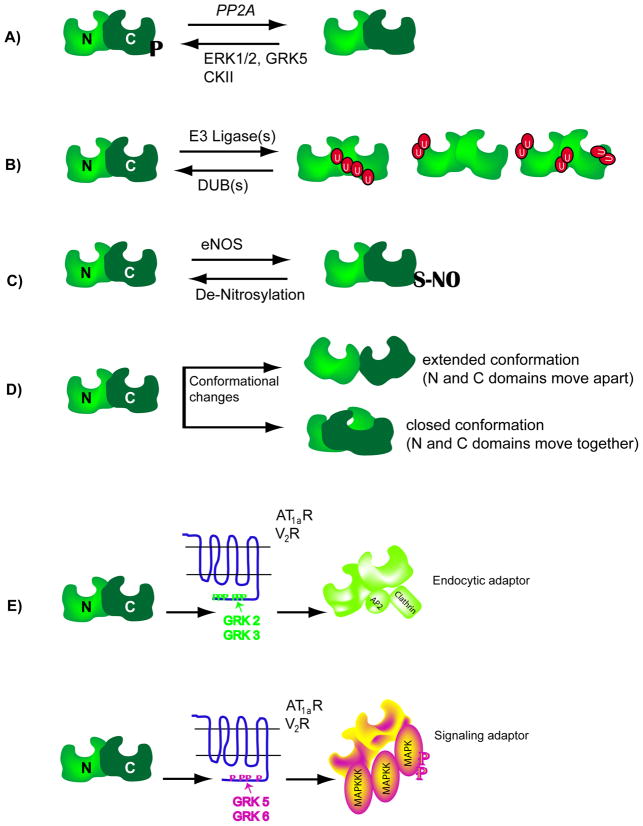
β-Arrestins function as endocytic adaptors and mediate trafficking of a variety of cell-surface receptors, including seven-transmembrane receptors (7TMRs). In the case of 7TMRs, β-arrestins carry out these tasks while simultaneously inhibiting upstream G-protein-dependent signaling and promoting alternate downstream signaling pathways. The mechanisms by which β-arrestins interact with a continuously expanding ensemble of protein partners and perform their multiple functions including trafficking and signaling are currently being uncovered. Molecular changes at the level of protein conformation as well as post-translational modifications of β-arrestins probably form the basis for their dynamic interactions during receptor trafficking and signaling. It is becoming increasingly evident that β-arrestins, originally discovered as 7TMR adaptor proteins, indeed have much broader and more versatile roles in maintaining cellular homeostasis. In this review paper, we assess the traditional and novel functions of β-arrestins and discuss the molecular attributes that might facilitate multiple interactions in regulating cell signaling and receptor trafficking.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures

References
-
- Lefkowitz RJ. Historical review: a brief history and personal retrospective of seven-transmembrane receptors. Trends Pharmacol Sci. 2004;25 (8):413–422. - PubMed
-
- Yamaki K, et al. Molecular cloning of the S-antigen cDNA from bovine retina. Biochem Biophys Res Commun. 1987;142 (3):904–910. - PubMed
-
- Lohse MJ, et al. beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science. 1990;248 (4962):1547–1550. - PubMed
-
- Attramadal H, et al. Beta-arrestin2, a novel member of the arrestin/beta-arrestin gene family. J Biol Chem. 1992;267 (25):17882–17890. - PubMed
-
- Smith WC, et al. Cloning and functional characterization of salamander rod and cone arrestins. Invest Ophthalmol Vis Sci. 2000;41 (9):2445–2455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
