

Perisomatic inhibition and cortical circuit dysfunction in schizophrenia
- PMID: 21680173
- PMCID: PMC3183273
- DOI: 10.1016/j.conb.2011.05.013
Perisomatic inhibition and cortical circuit dysfunction in schizophrenia
Abstract
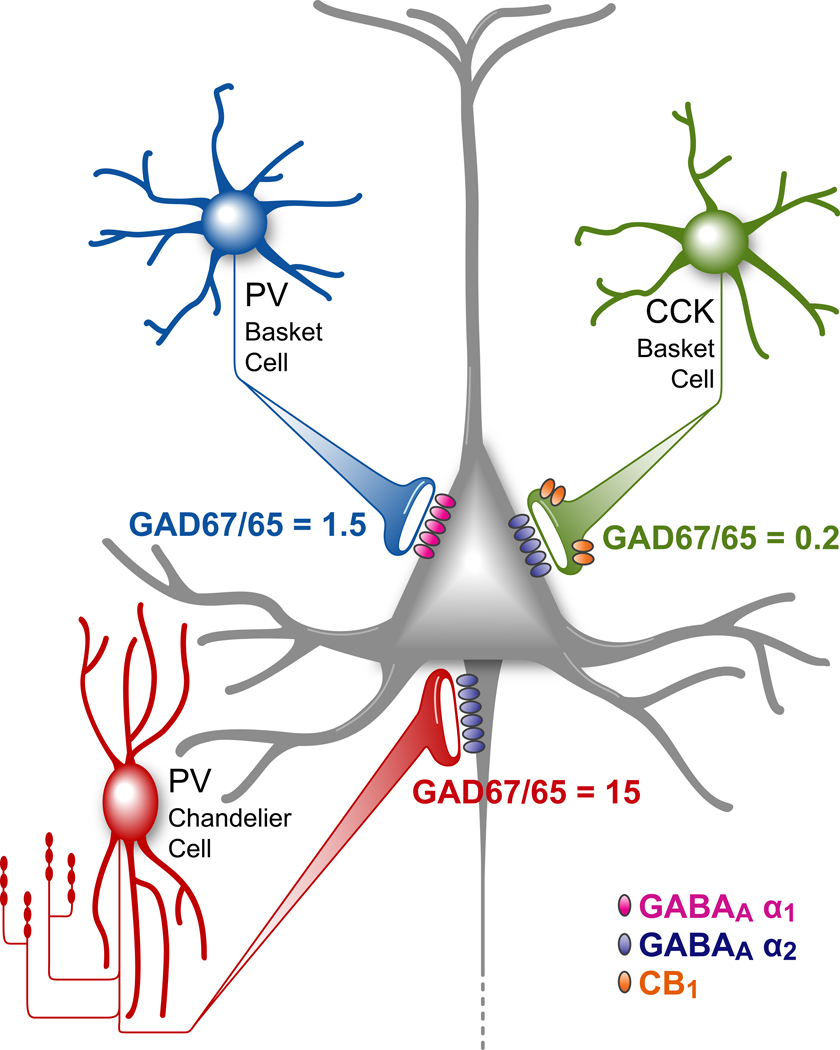
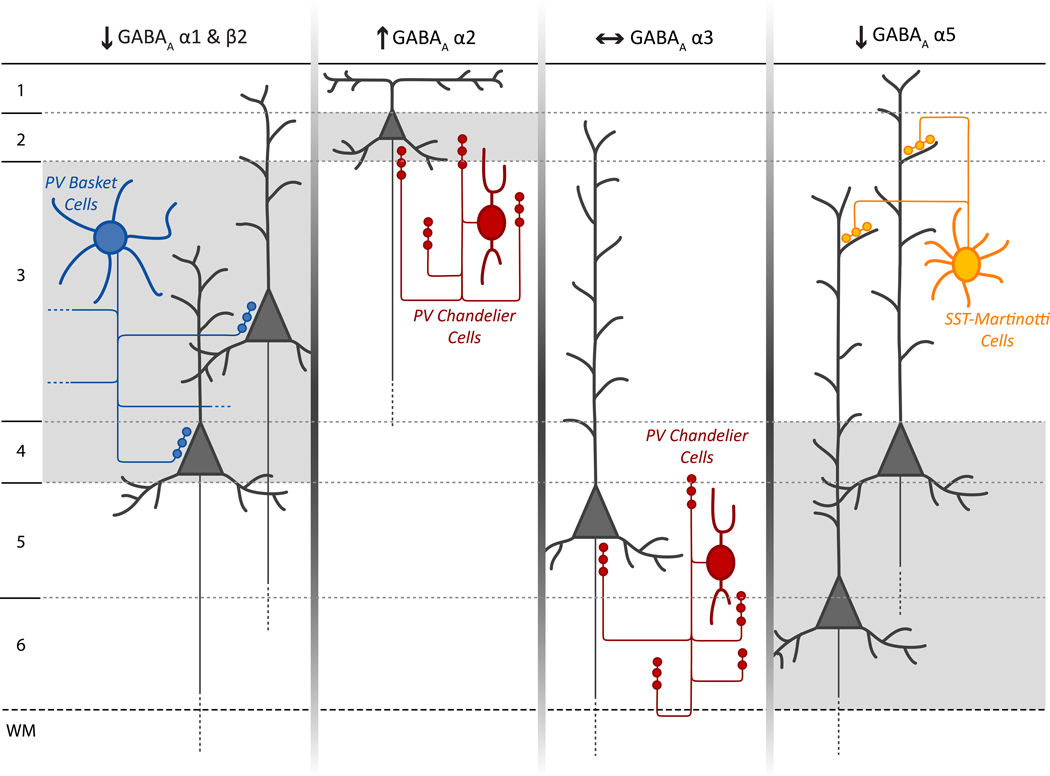
Deficits of cognitive control in schizophrenia are associated with altered gamma oscillations in the prefrontal cortex. Paralbumin basket interneurons, which innervate the perisomatic region of pyramidal neurons, appear to play a key role in generating cortical gamma oscillations. In the prefrontal cortex of subjects with schizophrenia, alterations are present in both pre- and post-synaptic markers of the strength of GABA inputs from parvalbumin basket neurons to pyramidal neurons. These alterations may contribute to the neural substrate for impaired gamma oscillations in schizophrenia.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures

References
-
-
Reichenberg A, Caspi A, Harrington H, Houts R, Keefe RS, Murray RM, Poulton R, Moffitt TE. Static and dynamic cognitive deficits in childhood preceding adult schizophrenia: a 30-year study. Am J Psychiatry. 2010;167:160–169. ** This study of the cognitive development of more than over 1,000 children over a 30-year period demonstrated that in individuals who later develop schizophrenia, some cognitive deficits are present at an early age and others are progressive before the onset of clinical symptoms.
-
-
-
Minzenberg MJ, Laird AR, Thelen S, Carter CS, Glahn DC. Meta-analysis of 41 functional neuroimaging studies of executive function in schizophrenia. Arch Gen Psychiatry. 2009;66:811–822. * An important meta-analysis of a large body of literature documenting altered functional activation of the dorsolateral prefrontal cortex during cognitive tasks in individuals with schizophrenia.
-
-
-
Minzenberg MJ, Firl AJ, Yoon JH, Gomes GC, Reinking C, Carter CS. Gamma oscillatory power is impaired during cognitive control independent of medication status in first-episode schizophrenia. Neuropsychopharm. 2010;35:2590–2599. * An important study showing that cognitive deficits and altered gamma oscillations are present in schizophrenia independent of medication effects or illness chronicity.
-
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
