

Contrasting 5' and 3' evolutionary histories and frequent evolutionary convergence in Meis/hth gene structures
- PMID: 21680890
- PMCID: PMC3140891
- DOI: 10.1093/gbe/evr056
Contrasting 5' and 3' evolutionary histories and frequent evolutionary convergence in Meis/hth gene structures
Abstract
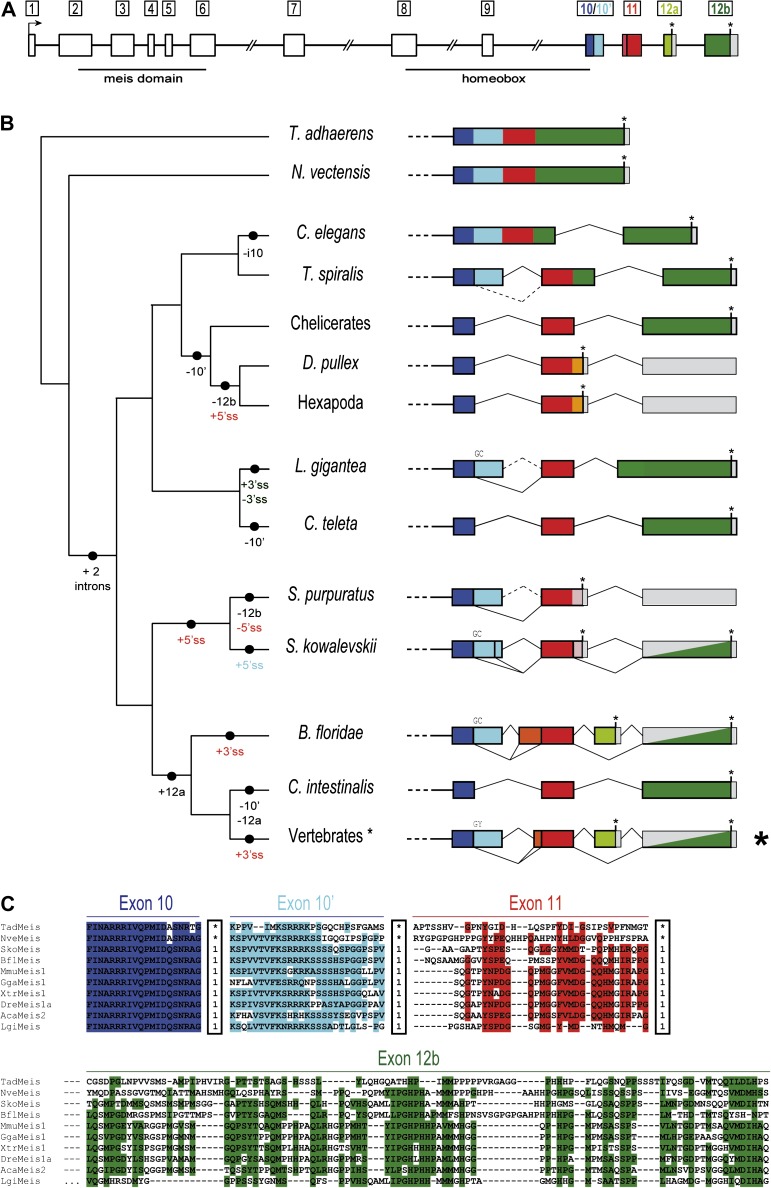
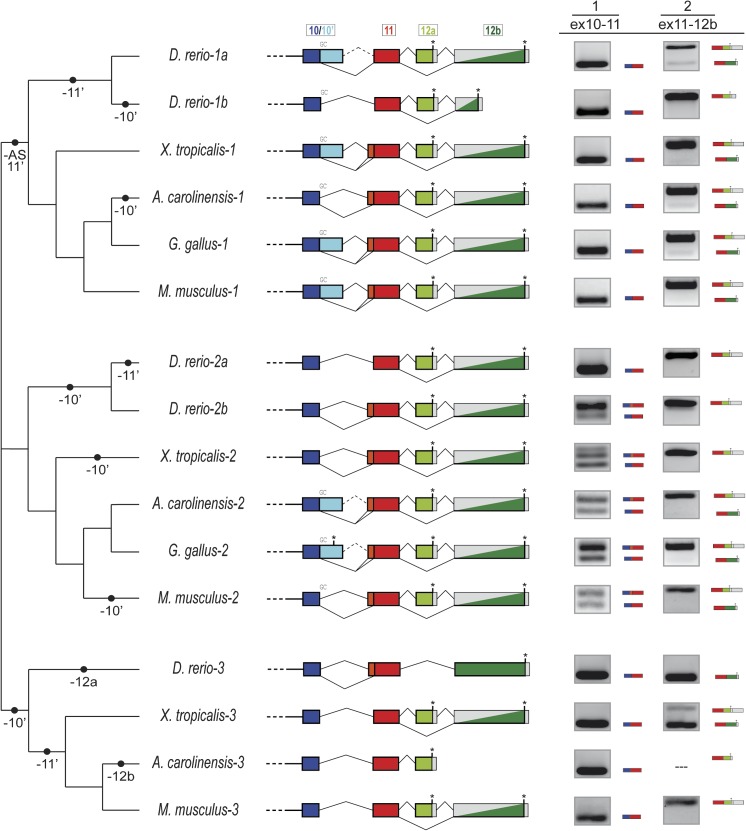
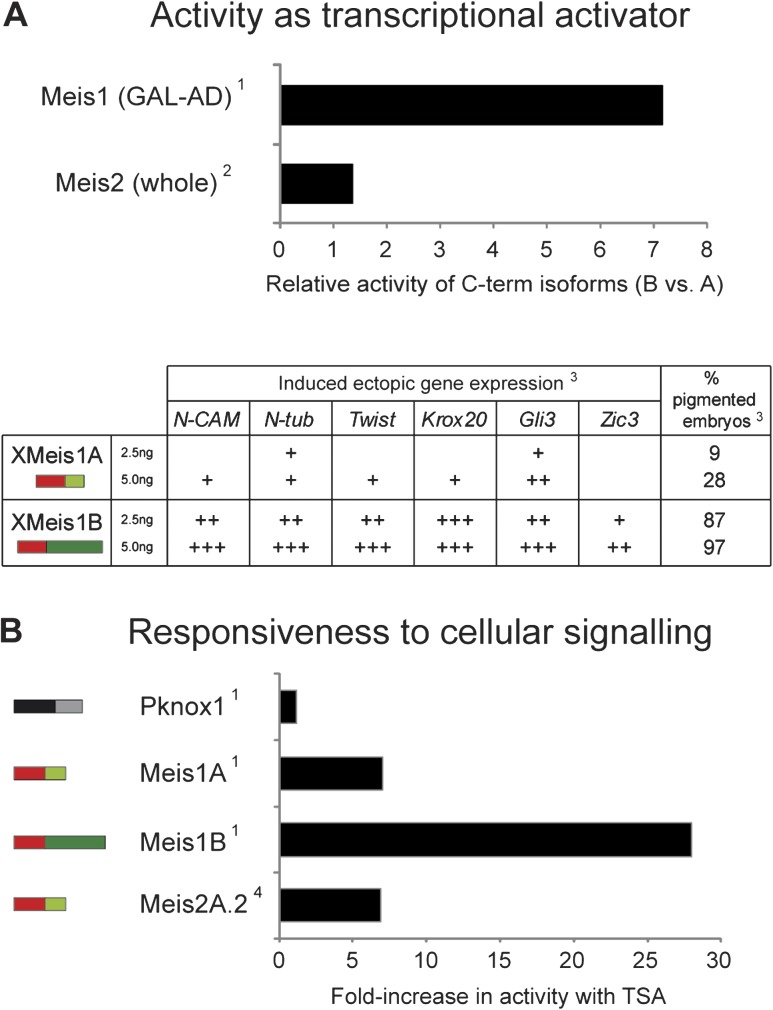
Organisms show striking differences in genome structure; however, the functional implications and fundamental forces that govern these differences remain obscure. The intron-exon organization of nuclear genes is involved in a particularly large variety of structures and functional roles. We performed a 22-species study of Meis/hth genes, intron-rich homeodomain-containing transcription factors involved in a wide range of developmental processes. Our study revealed three surprising results that suggest important and very different functions for Meis intron-exon structures. First, we find unexpected conservation across species of intron positions and lengths along most of the Meis locus. This contrasts with the high degree of structural divergence found in genome-wide studies and may attest to conserved regulatory elements residing within these conserved introns. Second, we find very different evolutionary histories for the 5' and 3' regions of the gene. The 5'-most 10 exons, which encode the highly conserved Meis domain and homeodomain, show striking conservation. By contrast, the 3' of the gene, which encodes several domains implicated in transcriptional activation and response to cell signaling, shows a remarkably active evolutionary history, with diverse isoforms and frequent creation and loss of new exons and splice sites. This region-specific diversity suggests evolutionary "tinkering," with alternative splicing allowing for more subtle regulation of protein function. Third, we find a large number of cases of convergent evolution in the 3' region, including 1) parallel losses of ancestral coding sequence, 2) parallel gains of external and internal splice sites, and 3) recurrent truncation of C-terminal coding regions. These results attest to the importance of locus-specific splicing functions in differences in structural evolution across genes, as well as to commonalities of forces shaping the evolution of individual genes along different lineages.
Figures
References
-
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005;21:2104–2105. - PubMed
-
- Adams MD, et al. The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. - PubMed
-
- Alexander RP, Fang G, Rozowsky J, Snyder M, Gerstein MB. Annotating non-coding regions of the genome. Nat Rev Genet. 2010;11:559–571. - PubMed
-
- Aparicio S, et al. Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science. 2002;297:1301–1310. - PubMed
-
- Azcoitia V, Aracil M, Martínez-A C, Torres M. The homeodomain protein Meis1 is essential for definitive hematopoiesis and vascular patterning in the mouse embryo. Dev Biol. 2005;280:307–320. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
