

RAGE-dependent activation of the oncoprotein Pim1 plays a critical role in systemic vascular remodeling processes
- PMID: 21680901
- PMCID: PMC3545710
- DOI: 10.1161/ATVBAHA.111.230573
RAGE-dependent activation of the oncoprotein Pim1 plays a critical role in systemic vascular remodeling processes
Abstract
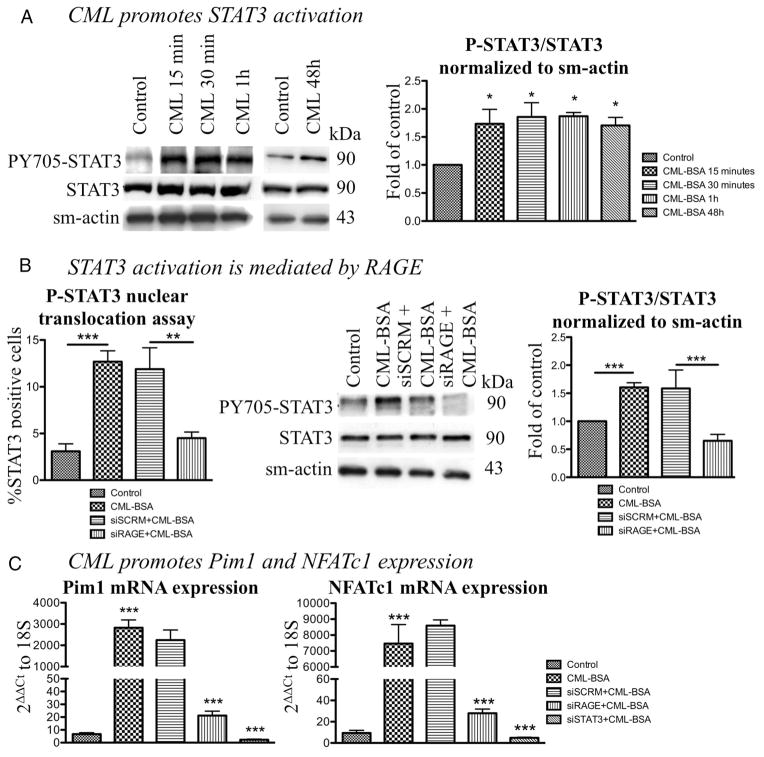
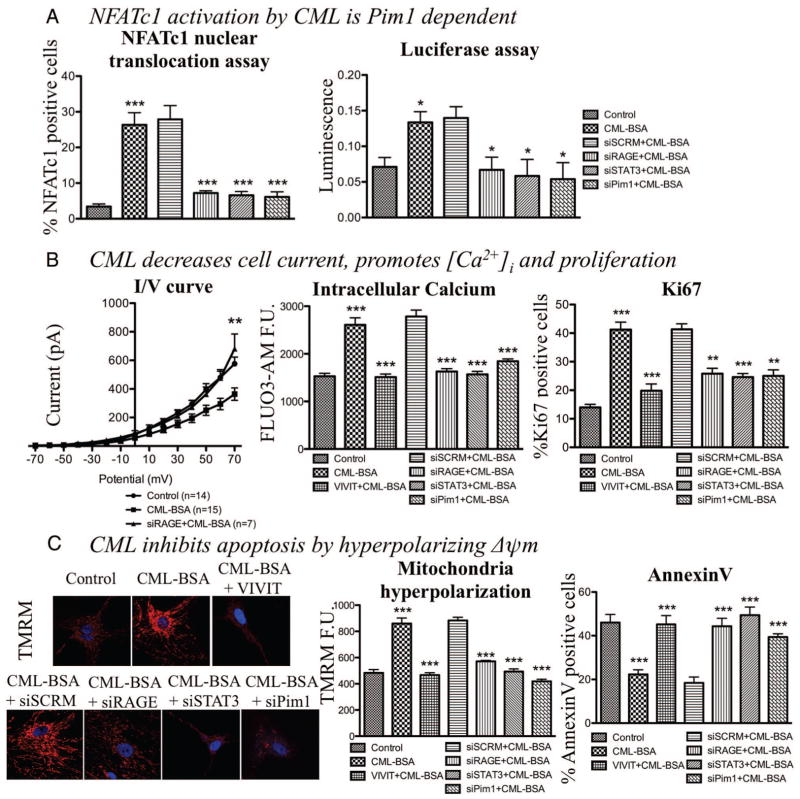
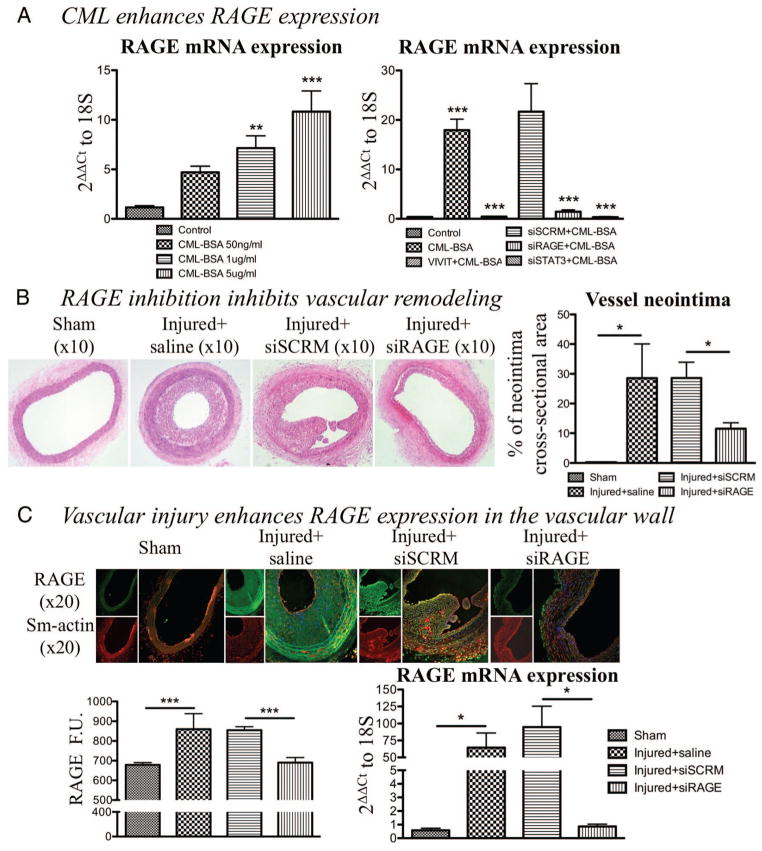
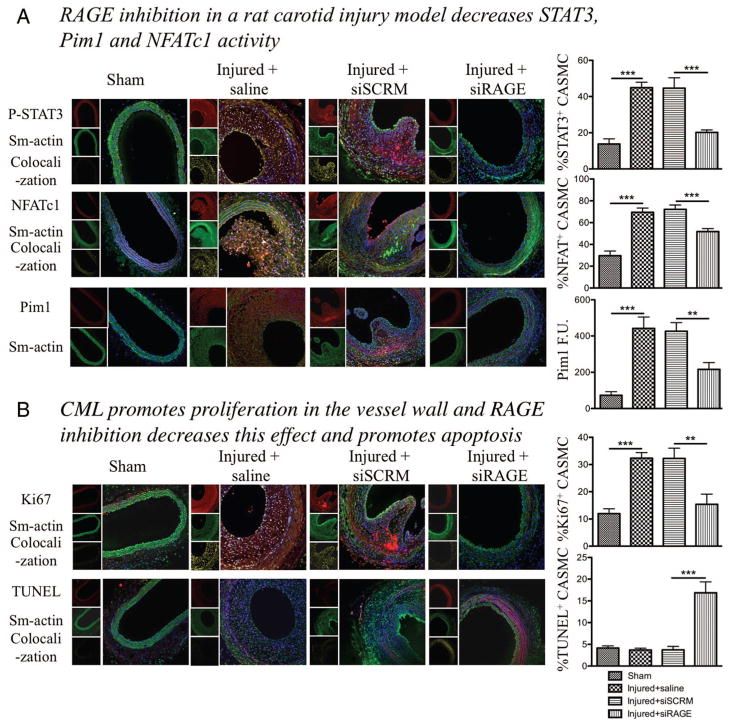
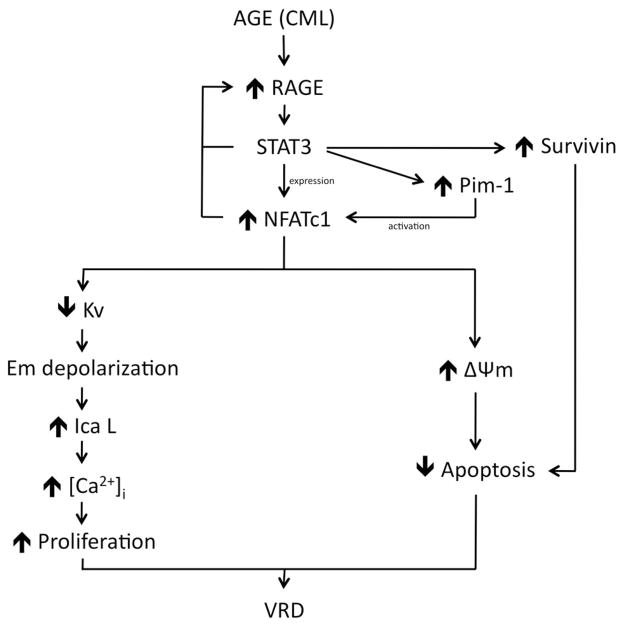
Objective: Vascular remodeling diseases (VRD) are mainly characterized by inflammation and a vascular smooth muscle cells (VSMCs) proproliferative and anti-apoptotic phenotype. Recently, the activation of the advanced glycation endproducts receptor (RAGE) has been shown to promote VSMC proliferation and resistance to apoptosis in VRD in a signal transducer and activator of transcription (STAT)3-dependant manner. Interestingly, we previously described in both cancer and VRD that the sustainability of this proproliferative and antiapoptotic phenotype requires activation of the transcription factor NFAT (nuclear factor of activated T-cells). In cancer, NFAT activation is dependent of the oncoprotein provirus integration site for Moloney murine leukemia virus (Pim1), which is regulated by STAT3 and activated in VRD. Therefore, we hypothesized that RAGE/STAT3 activation in VSMC activates Pim1, promoting NFAT and thus VSMC proliferation and resistance to apoptosis. Methods/Results- In vitro, freshly isolated human carotid VSMCs exposed to RAGE activator Nε-(carboxymethyl)lysine (CML) for 48 hours had (1) activated STAT3 (increased P-STAT3/STAT3 ratio and P-STAT3 nuclear translocation); (2) increased STAT3-dependent Pim1 expression resulting in NFATc1 activation; and (3) increased Pim1/NFAT-dependent VSMC proliferation (PCNA, Ki67) and resistance to mitochondrial-dependent apoptosis (TMRM, Annexin V, TUNEL). Similarly to RAGE inhibition (small interfering RNA [siRNA]), Pim1, STAT3 and NFATc1 inhibition (siRNA) reversed these abnormalities in human carotid VSMC. Moreover, carotid artery VSMCs isolated from Pim1 knockout mice were resistant to CML-induced VSMC proliferation and resistance to apoptosis. In vivo, RAGE inhibition decreases STAT3/Pim1/NFAT activation, reversing vascular remodeling in the rat carotid artery-injured model.
Conclusions: RAGE activation accounts for many features of VRD including VSMC proliferation and resistance to apoptosis by the activation of STAT3/Pim1/NFAT axis. Molecules aimed to inhibit RAGE could be of a great therapeutic interest for the treatment of VRD.
Figures
References
-
- Bonnet S, Paulin R, Sutendra G, Dromparis P, Roy M, Watson KO, Nagendran J, Haromy A, Dyck JR, Michelakis ED. Dehydroepiandrosterone reverses systemic vascular remodeling through the inhibition of the Akt/GSK3-{beta}/NFAT axis. Circulation. 2009;120:1231–1240. - PubMed
-
- Lambert CM, Roy M, Robitaille GA, Richard DE, Bonnet S. HIF-1 inhibition decreases systemic vascular remodelling diseases by promoting apoptosis through a hexokinase 2-dependent mechanism. Cardiovasc Res. 88:196–204. - PubMed
-
- Libby P, Clinton SK. Cytokines as mediators of vascular pathology. Nouv Rev Fr Hematol. 1992;34 (Suppl):S47–S53. - PubMed
-
- Schwartz LB, Radic ZS, O’Donohoe MK, Mikat EM, McCann RL, Hagen PO. Saphenous vein endothelium-dependent relaxation in patients with peripheral vascular disease. Ann Vasc Surg. 1992;6:425– 432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
