

Smad6 is essential to limit BMP signaling during cartilage development
- PMID: 21681813
- PMCID: PMC3183270
- DOI: 10.1002/jbmr.443
Smad6 is essential to limit BMP signaling during cartilage development
Abstract
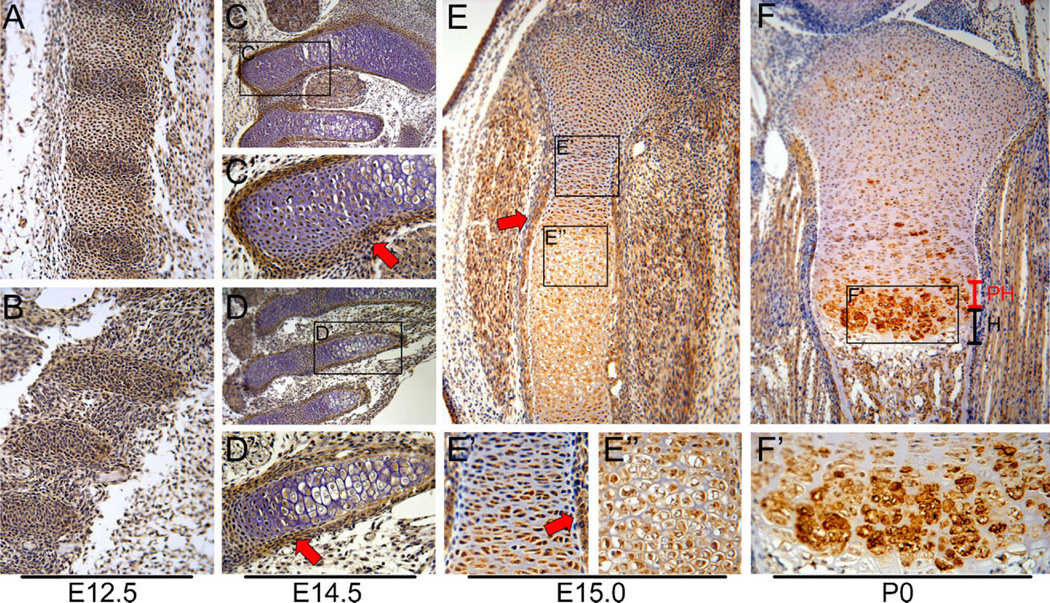
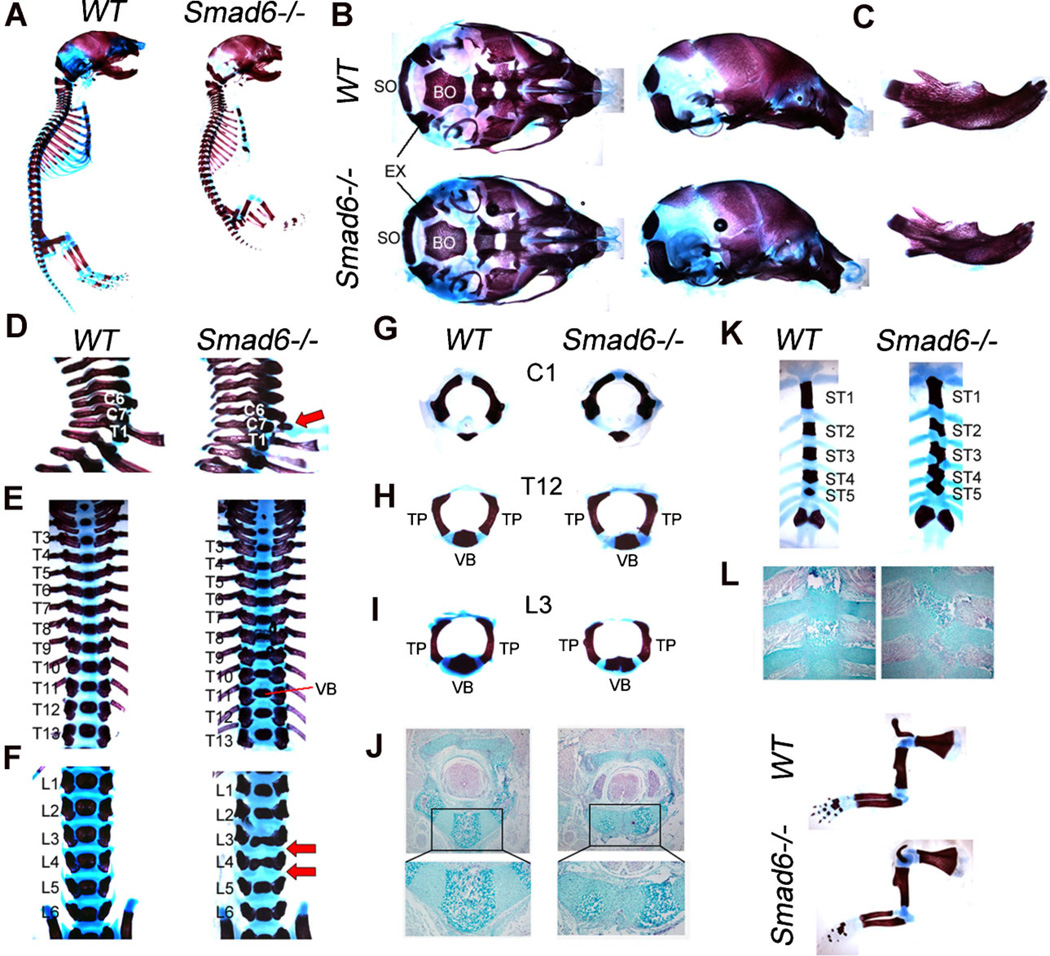
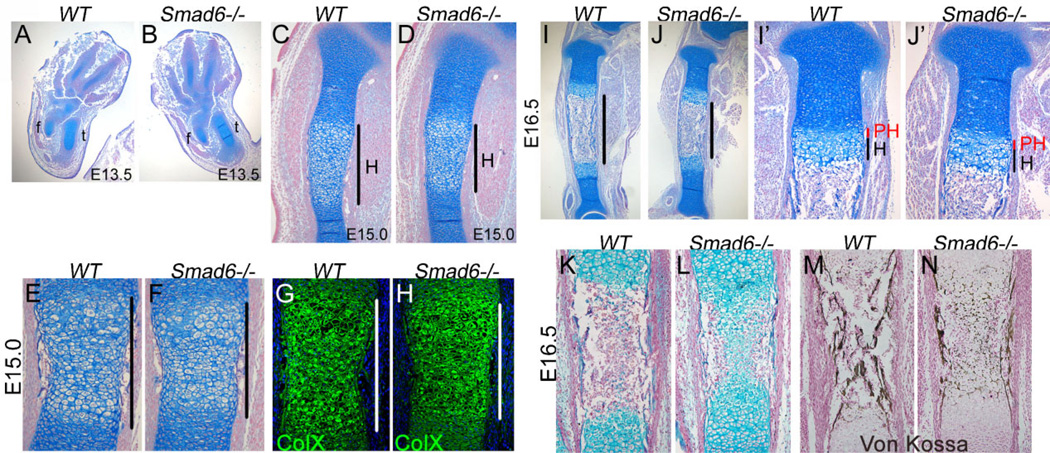
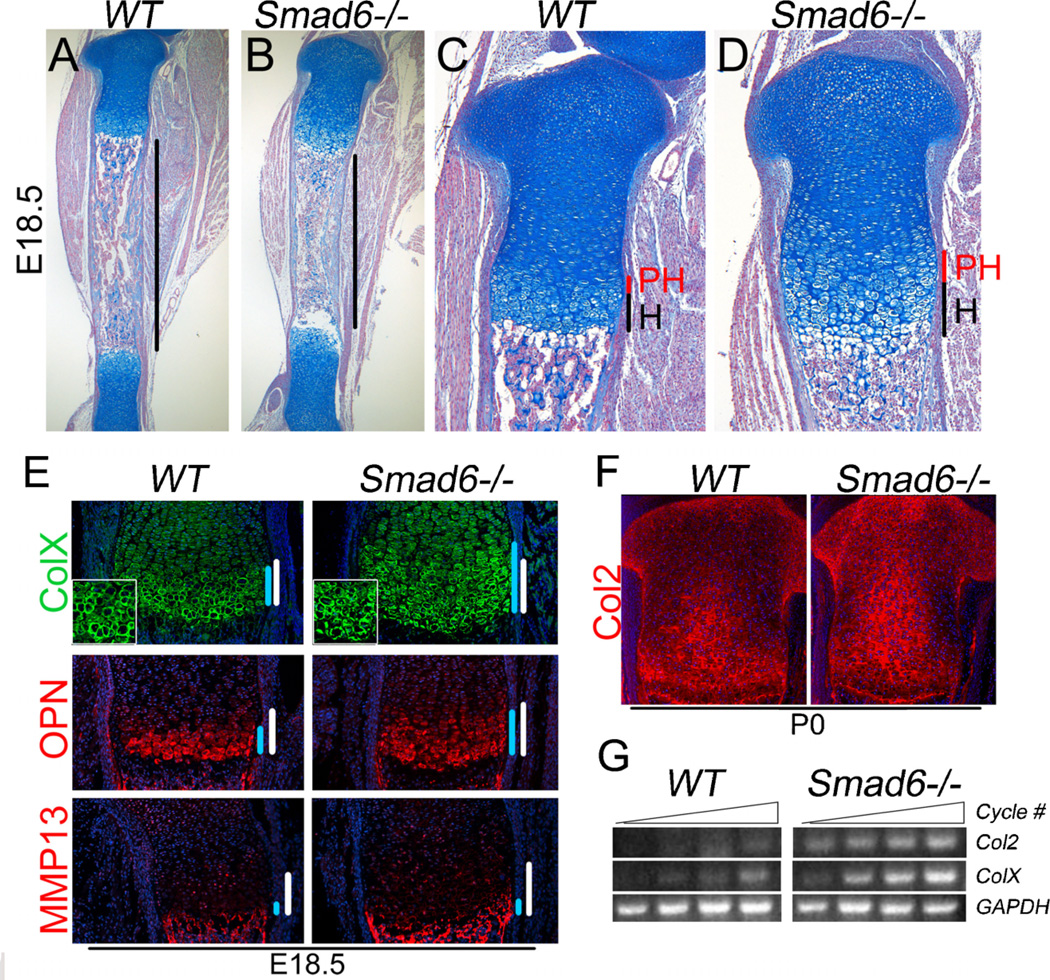
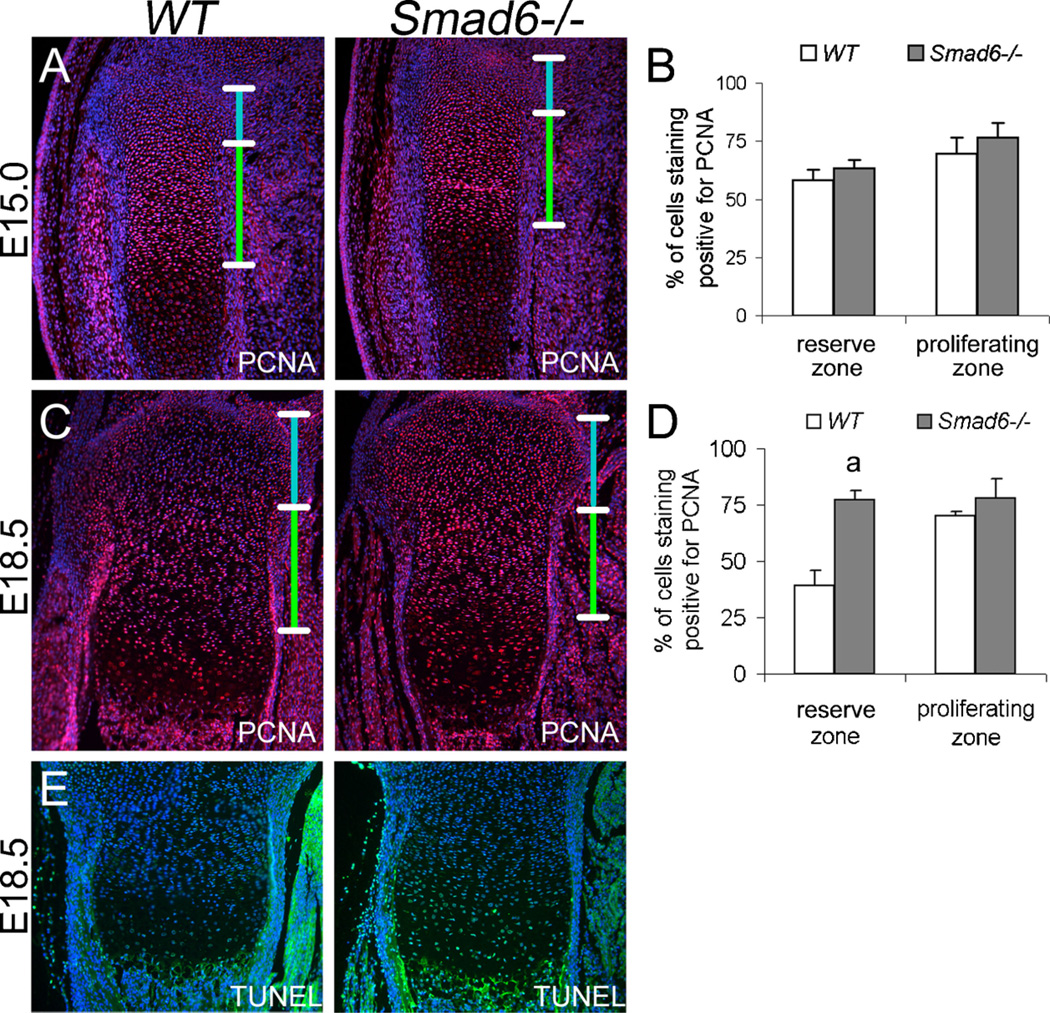
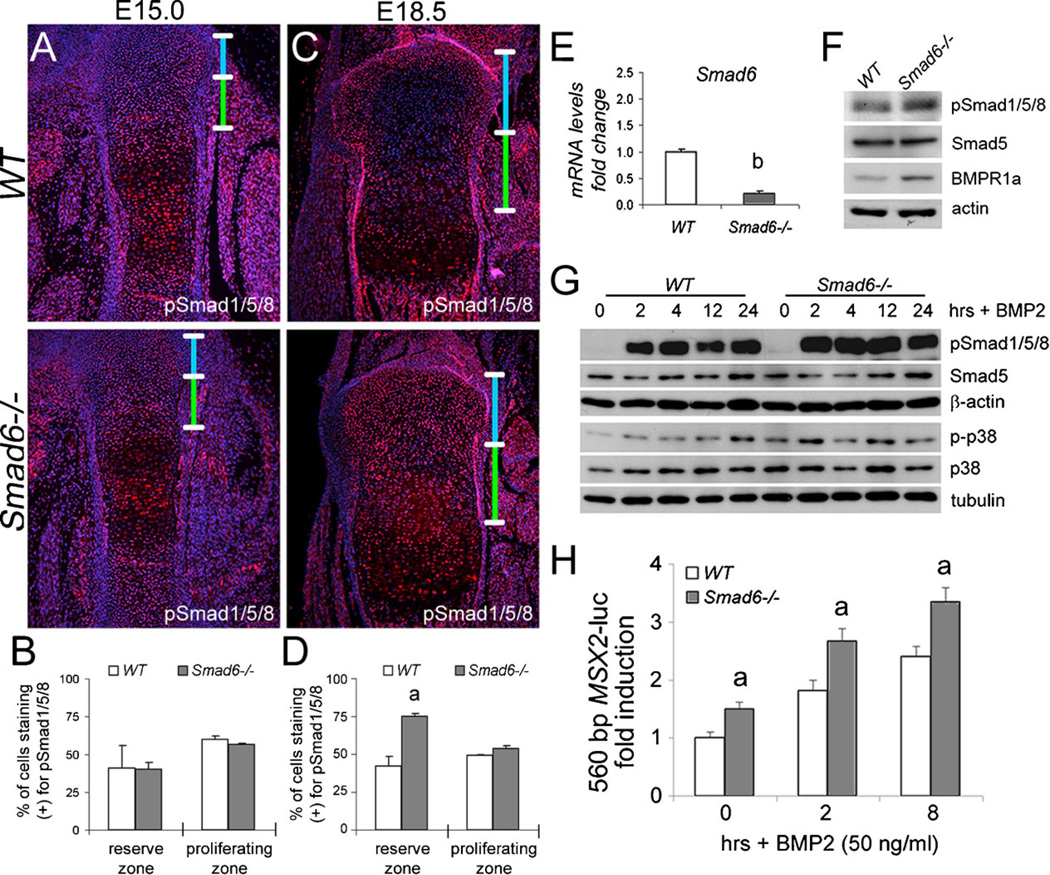
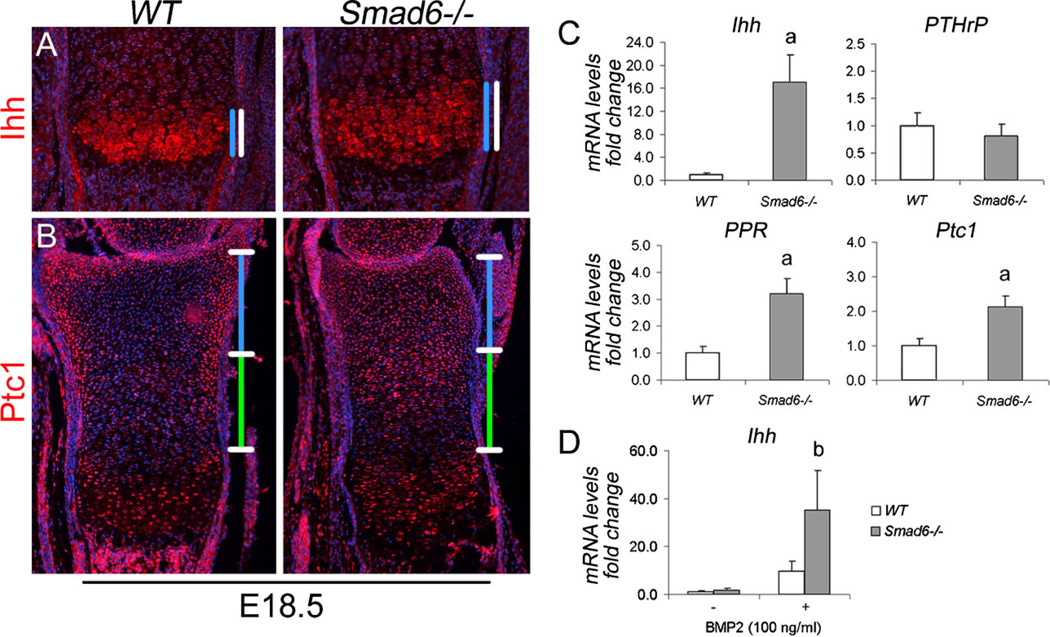
Bone morphogenetic protein (BMP) signaling pathways regulate multiple aspects of endochondral bone formation. The importance of extracellular antagonists as regulators of BMP signaling has been defined. In vitro studies reveal that the intracellular regulators, inhibitory Smads 6 and 7, can regulate BMP-mediated effects on chondrocytes. Although in vivo studies in which inhibitory Smads were overexpressed in cartilage have shown that inhibitory Smads have the potential to limit BMP signaling in vivo, the physiological relevance of inhibitory Smad activity in skeletal tissues is unknown. In this study, we have determined the role of Smad6 in endochondral bone formation. Loss of Smad6 in mice leads to defects in both axial and appendicular skeletal development. Specifically, Smad6-/- mice exhibit a posterior transformation of the seventh cervical vertebra, bilateral ossification centers in lumbar vertebrae, and bifid sternebrae due to incomplete sternal band fusion. Histological analysis of appendicular bones revealed delayed onset of hypertrophic differentiation and mineralization at midgestation in Smad6-/- mice. By late gestation, however, an expanded hypertrophic zone, associated with an increased pool of proliferating cells undergoing hypertrophy, was evident in Smad6 mutant growth plates. The mutant phenotype is attributed, at least in part, to increased BMP responsiveness in Smad6-deficient chondrocytes. Overall, our results show that Smad6 is required to limit BMP signaling during endochondral bone formation.
Copyright © 2011 American Society for Bone and Mineral Research.
Conflict of interest statement
Figures
References
-
- Kronenberg HM. Developmental regulation of the growth plate. Nature. 2003;423(6937):332–336. - PubMed
-
- Zehentner BK, Dony C, Burtscher H. The transcription factor Sox9 is involved in BMP-2 signaling. J Bone Miner Res. 1999;14(10):1734–1741. - PubMed
-
- Pan Q, Yu Y, Chen Q, Li C, Wu H, Wan Y, Ma J, Sun F. Sox9, a key transcription factor of bone morphogenetic protein-2-induced chondrogenesis, is activated through BMP pathway and a CCAAT box in the proximal promoter. J Cell Physiol. 2008;217(1):228–241. - PubMed
-
- Valcourt U, Gouttenoire J, Moustakas A, Herbage D, Mallein-Gerin F. Functions of transforming growth factor-beta family type I receptors and Smad proteins in the hypertrophic maturation and osteoblastic differentiation of chondrocytes. J Biol Chem. 2002;277(37):33545–33558. - PubMed
-
- Minina E, Wenzel HM, Kreschel C, Karp S, Gaffield W, McMahon AP, Vortkamp A. BMP and Ihh/PTHrP signaling interact to coordinate chondrocyte proliferation and differentiation. Development. 2001;128(22):4523–4534. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
