

Activation of natural killer cells by hepatitis C virus particles in vitro
- PMID: 21682720
- PMCID: PMC3170984
- DOI: 10.1111/j.1365-2249.2011.04431.x
Activation of natural killer cells by hepatitis C virus particles in vitro
Abstract
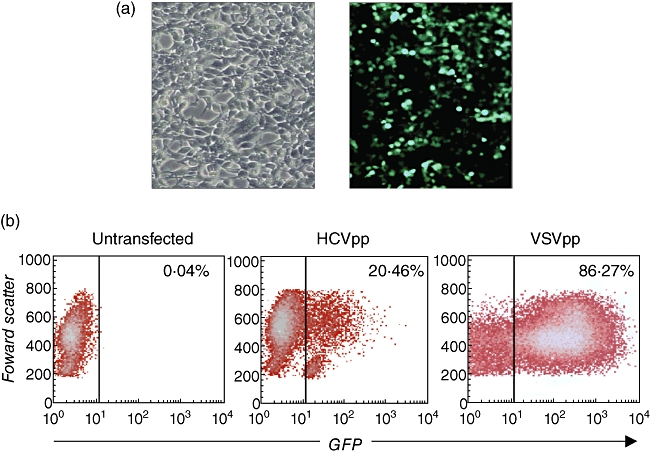
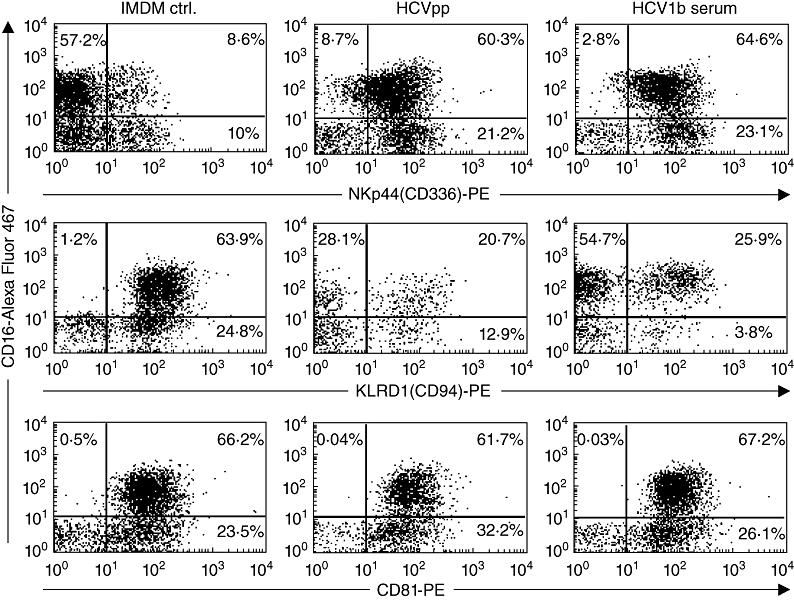
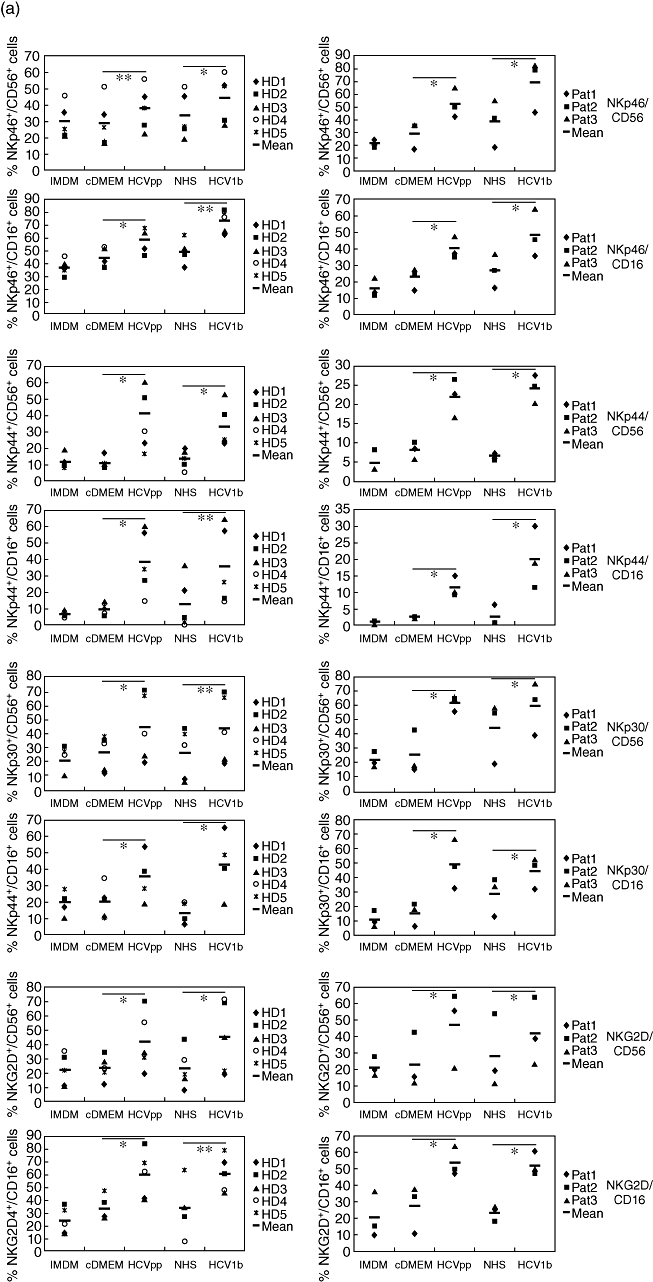
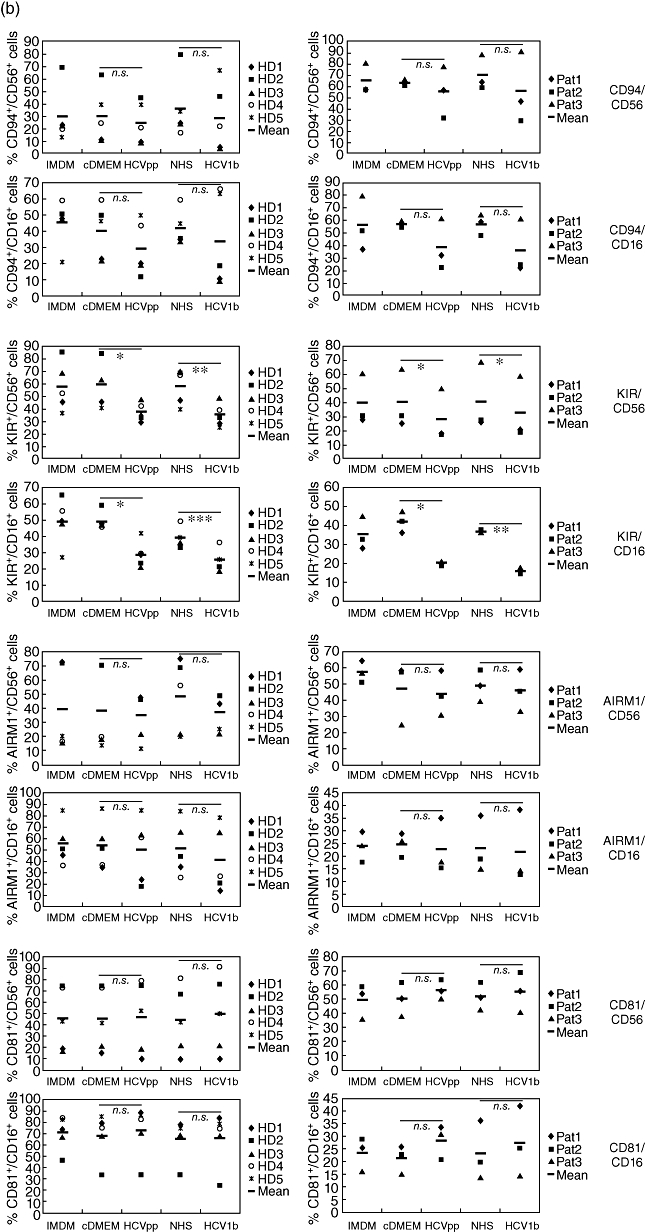
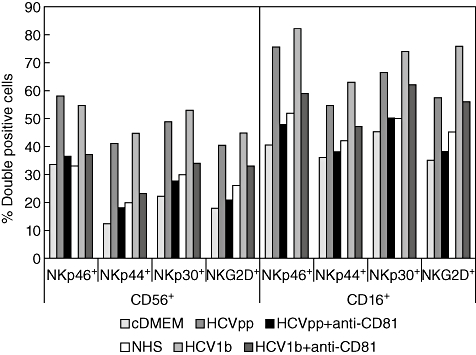
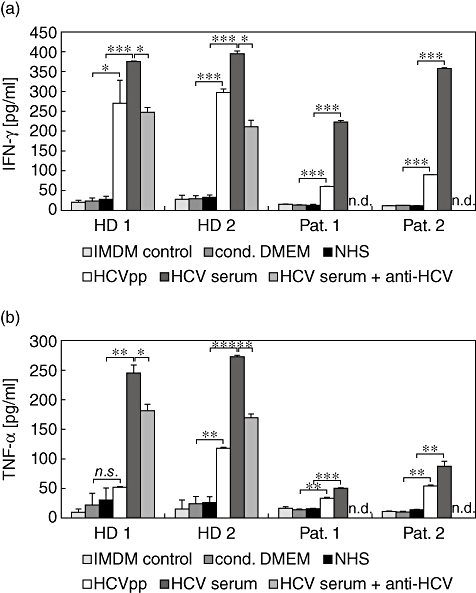
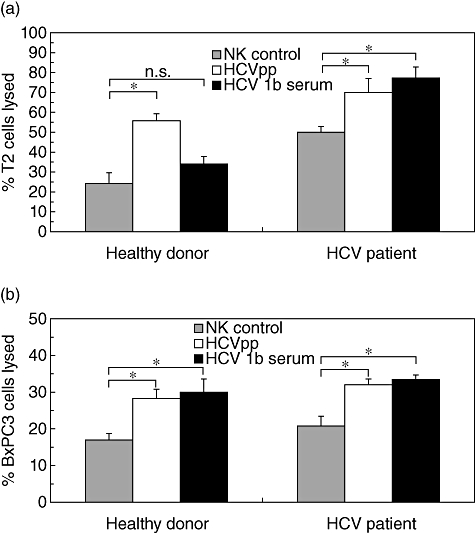
Little is known about the ability of hepatitis C virus (HCV) to alter early innate immune responses in infected patients. Previous studies have shown that natural killer (NK) cells are functionally impaired after interaction of recombinant HCV glycoprotein E2 with the co-stimulatory CD81 molecule in vitro; however, the functional consequences of a prolonged contact of NK cells with HCV particles have remained unclear. We have examined the phenotypes of purified, interleukin-2-activated NK cells from healthy donors and HCV genotype 1b patients after culture for 5 days with HCV pseudoparticles (HCVpp) and serum samples containing HCV genotype 1b. NK cells from healthy donors and chronic HCV patients were found to up-regulate receptors associated with activation (NKp46, NKp44, NKp30, NKG2D), while NK receptors from the killer cell immunoglobulin-like receptor family (KIR/CD158), predominantly having an inhibitory function, were significantly down-modulated after culture in the presence of HCV particles compared with control cultures of NK cells. HCV-infected sera and HCVpp elicited significantly higher secretion of the NK effector lymphokines interferon-γ and tumour necrosis factor-α. Furthermore, HCV stimulated the cytotoxic potential of NK cells from normal donors and patients. The enhanced activation of NK cells after prolonged culture with HCVpp or HCV-containing sera for 5 days suggests that these innate effector cells may play an important role in viral control during early phases of HCV infection.
© 2011 The Authors. Clinical and Experimental Immunology © 2011 British Society for Immunology.
Figures
Similar articles
-
Human natural killer cells expressing the memory-associated marker CD45RO from tuberculous pleurisy respond more strongly and rapidly than CD45RO- natural killer cells following stimulation with interleukin-12.Immunology. 2011 Sep;134(1):41-9. doi: 10.1111/j.1365-2567.2011.03464.x. Epub 2011 Jun 29. Immunology. 2011. PMID: 21711347 Free PMC article.
-
Activating NK cell receptor expression/function (NKp30, NKp46, DNAM-1) during chronic viraemic HCV infection is associated with the outcome of combined treatment.Eur J Immunol. 2011 Oct;41(10):2905-14. doi: 10.1002/eji.201041361. Eur J Immunol. 2011. PMID: 21695691
-
Reduced frequencies of NKp30+NKp46+, CD161+, and NKG2D+ NK cells in acute HCV infection may predict viral clearance.J Hepatol. 2011 Aug;55(2):278-88. doi: 10.1016/j.jhep.2010.11.030. Epub 2010 Dec 17. J Hepatol. 2011. PMID: 21168454 Free PMC article.
-
NK cells prevalence, subsets and function in viral hepatitis C.Arch Immunol Ther Exp (Warsz). 2011 Dec;59(6):449-55. doi: 10.1007/s00005-011-0145-y. Epub 2011 Oct 5. Arch Immunol Ther Exp (Warsz). 2011. PMID: 21972016 Review.
-
The Natural Cytotoxicity Receptors in Health and Disease.Front Immunol. 2019 May 7;10:909. doi: 10.3389/fimmu.2019.00909. eCollection 2019. Front Immunol. 2019. PMID: 31134055 Free PMC article. Review.
Cited by
-
CD81 and hepatitis C virus (HCV) infection.Viruses. 2014 Feb 6;6(2):535-72. doi: 10.3390/v6020535. Viruses. 2014. PMID: 24509809 Free PMC article. Review.
-
Virus-mediated inhibition of natural cytotoxicity receptor recognition.Cell Mol Life Sci. 2012 Dec;69(23):3911-20. doi: 10.1007/s00018-012-1001-x. Epub 2012 May 1. Cell Mol Life Sci. 2012. PMID: 22547090 Free PMC article. Review.
-
NK Cell-Mediated Antitumor Effects of a Folate-Conjugated Immunoglobulin Are Enhanced by Cytokines.Cancer Immunol Res. 2016 Apr;4(4):323-336. doi: 10.1158/2326-6066.CIR-15-0168. Epub 2016 Feb 10. Cancer Immunol Res. 2016. PMID: 26865456 Free PMC article.
-
The Evolutionary Arms Race between Virus and NK Cells: Diversity Enables Population-Level Virus Control.Viruses. 2019 Oct 17;11(10):959. doi: 10.3390/v11100959. Viruses. 2019. PMID: 31627371 Free PMC article. Review.
-
Chronic HCV infection affects the NK cell phenotype in the blood more than in the liver.PLoS One. 2014 Aug 22;9(8):e105950. doi: 10.1371/journal.pone.0105950. eCollection 2014. PLoS One. 2014. PMID: 25148254 Free PMC article.
References
-
- Rehermann B, Nascimbeni M. Immunology of hepatitis B virus and hepatitis C virus infection. Nat Rev Immunol. 2005;5:215–29. - PubMed
-
- Golden-Mason L, Rosen HR. Natural killer cells: primary target for hepatitis C virus immune evasion strategies. Liver Transpl. 2006;12:363–72. - PubMed
-
- Szabo G, Chang S, Dolganiuc A. Altered innate immunity in chronic hepatitis C infection – cause or effect? Hepatology. 2007;46:1279–90. - PubMed
-
- Li Y, Zhang T, Ho C, et al. Natural killer cells inhibit hepatitis C virus expression. J Leukoc Biol. 2004;76:1171–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
