

Osmolyte effects on protein stability and solubility: a balancing act between backbone and side-chains
- PMID: 21683504
- PMCID: PMC3166983
- DOI: 10.1016/j.bpc.2011.05.012
Osmolyte effects on protein stability and solubility: a balancing act between backbone and side-chains
Abstract
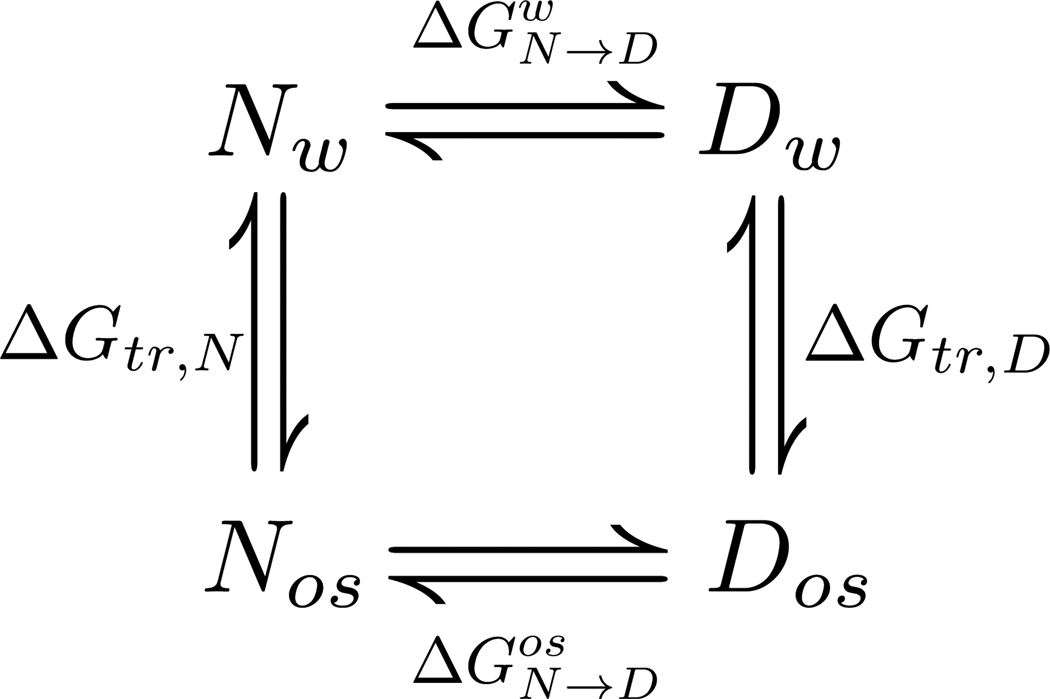
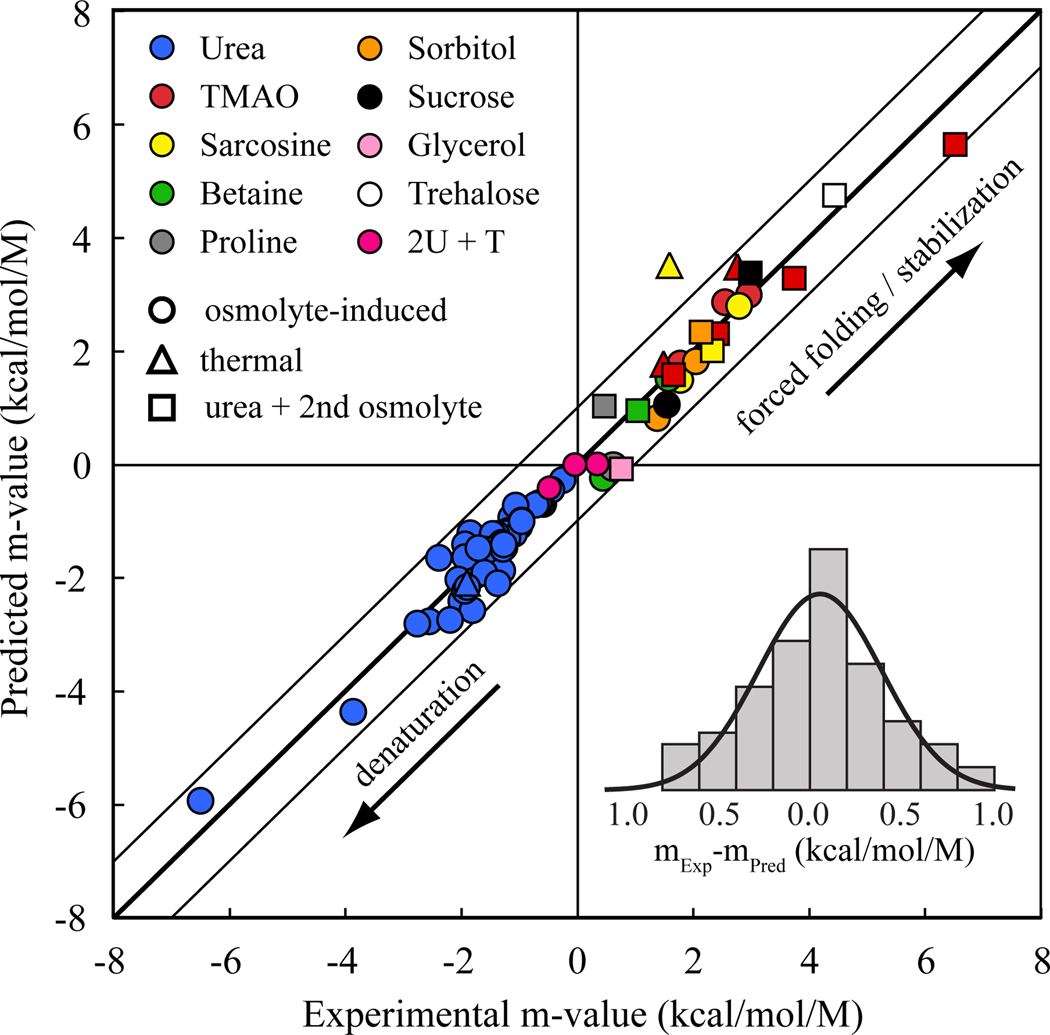
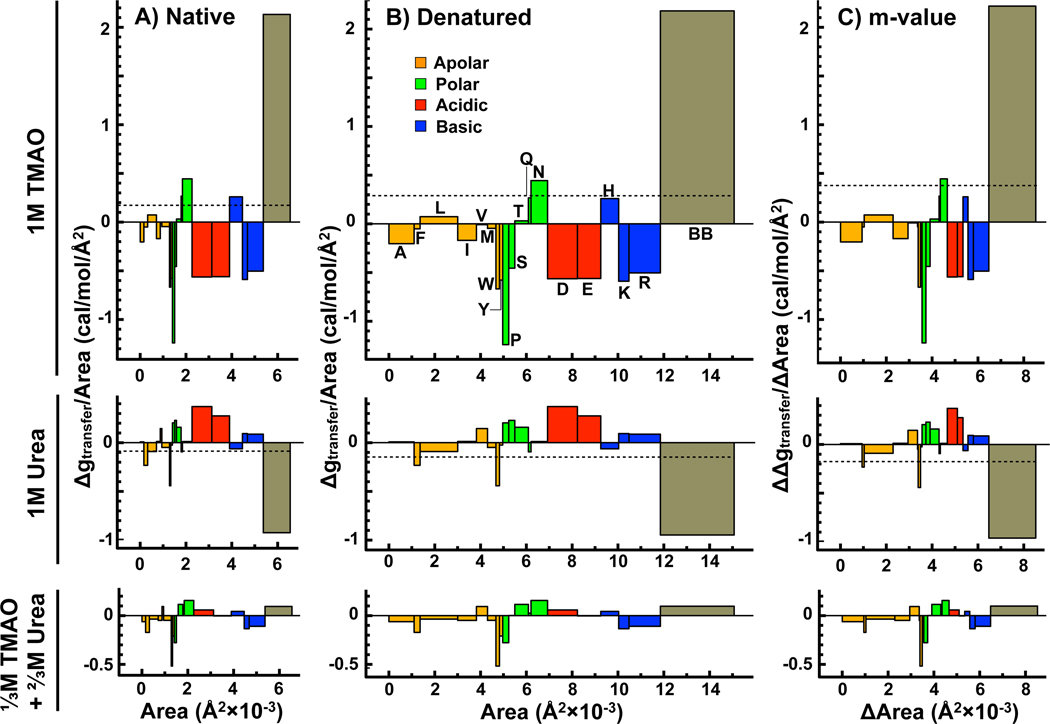
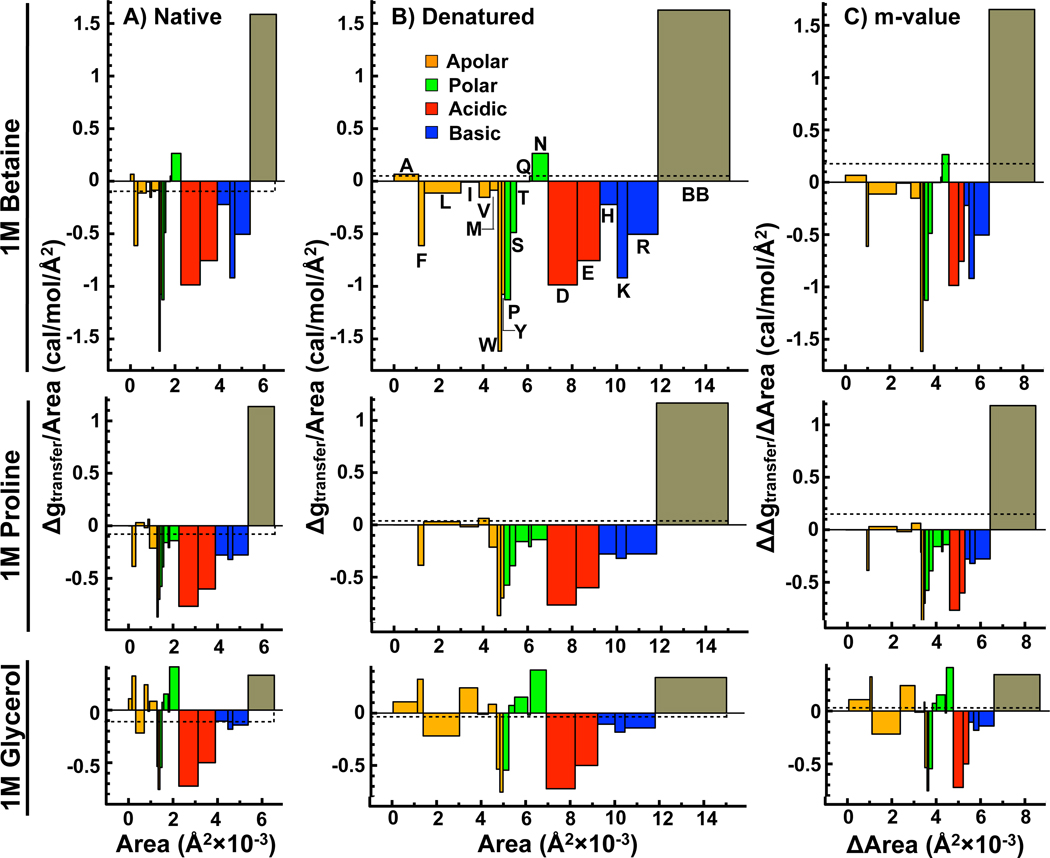
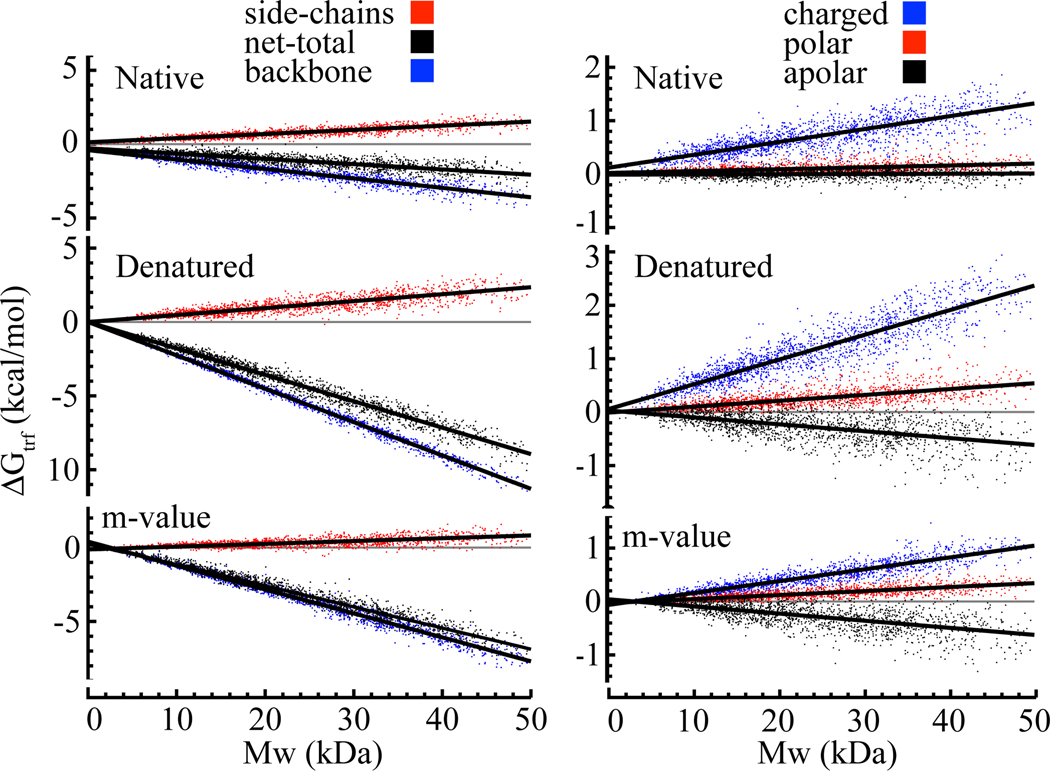
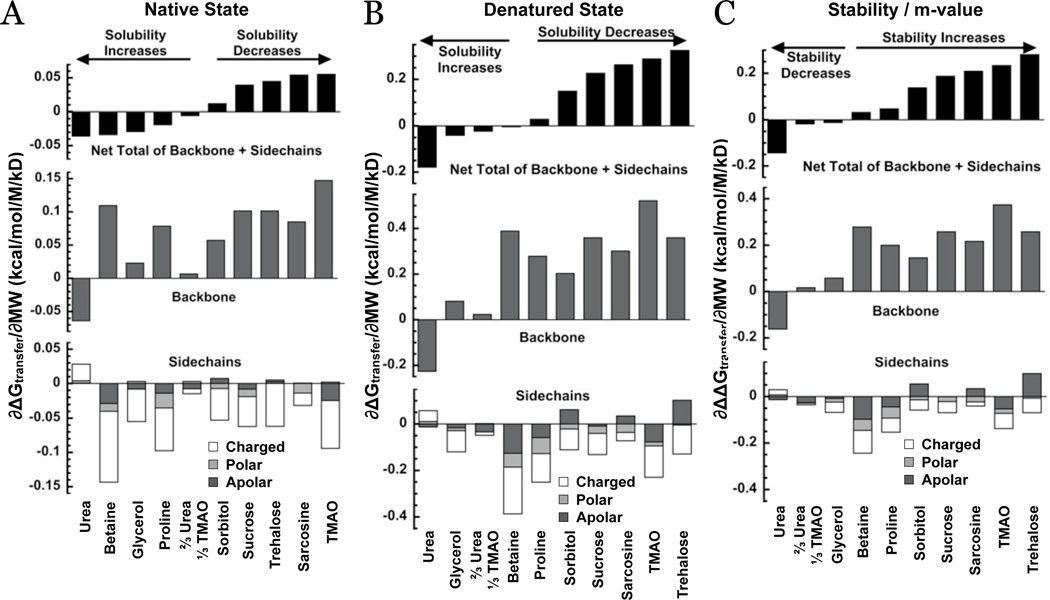
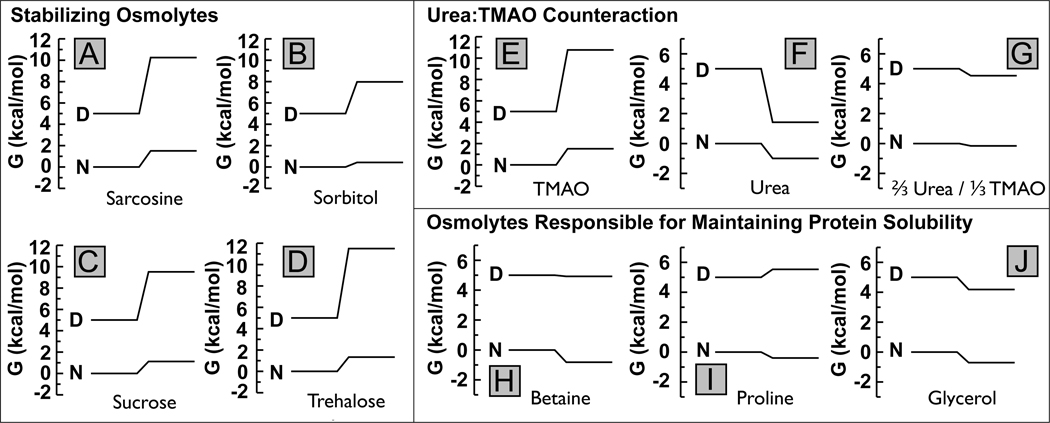
In adaptation biology the discovery of intracellular osmolyte molecules that in some cases reach molar levels, raises questions of how they influence protein thermodynamics. We've addressed such questions using the premise that from atomic coordinates, the transfer free energy of a native protein (ΔG(tr,N)) can be predicted by summing measured water-to-osmolyte transfer free energies of the protein's solvent exposed side chain and backbone component parts. ΔG(tr,D) is predicted using a self avoiding random coil model for the protein, and ΔG(tr,D)-ΔG(tr,N), predicts the m-value, a quantity that measures the osmolyte effect on the N⇌D transition. Using literature and newly measured m-values we show 1:1 correspondence between predicted and measured m-values covering a range of 12 kcal/mol/M in protein stability for 46 proteins and 9 different osmolytes. Osmolytes present a range of side chain and backbone effects on N and D solubility and protein stability key to their biological roles.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
References
-
- Hochachka P, Somero G. Mechanism and Process in Physiological Evolution. Oxford: Oxford University Press; 2002. Biochemical Adaptation.
-
- Yancey P, Clark M, Hand S, Bowlus R, Somero G. Living with water stress: Evolution of osmolyte systems. Science. 1982;217(4566):1214–1222. - PubMed
-
- Somero G. Protons, osmolytes, and fitness of internal milieu for protein function. Am. J. Physiol. 1986;251(2 Pt 2):R197–R213. - PubMed
-
- Gillett MB, Suko JR, Santoso FO, Yancey PH. Elevated levels of trimethylamine oxide in muscles of deep-sea gadiform teleosts: A high-pressure adaptation? J. Exp. Zool. 1997;279:386–391.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
