

Efficient computation via sparse coding in electrosensory neural networks
- PMID: 21683574
- PMCID: PMC4529320
- DOI: 10.1016/j.conb.2011.05.016
Efficient computation via sparse coding in electrosensory neural networks
Abstract
The electric sense combines spatial aspects of vision and touch with temporal features of audition. Its accessible neural architecture shares similarities with mammalian sensory systems and allows for recordings from successive brain areas to test hypotheses about neural coding. Further, electrosensory stimuli encountered during prey capture, navigation, and communication, can be readily synthesized in the laboratory. These features enable analyses of the neural circuitry that reveal general principles of encoding and decoding, such as segregation of information into separate streams and neural response sparsification. A systems level understanding arises via linkage between cellular differentiation and network architecture, revealed by in vitro and in vivo analyses, while computational modeling reveals how single cell dynamics and connectivity shape the sparsification process.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
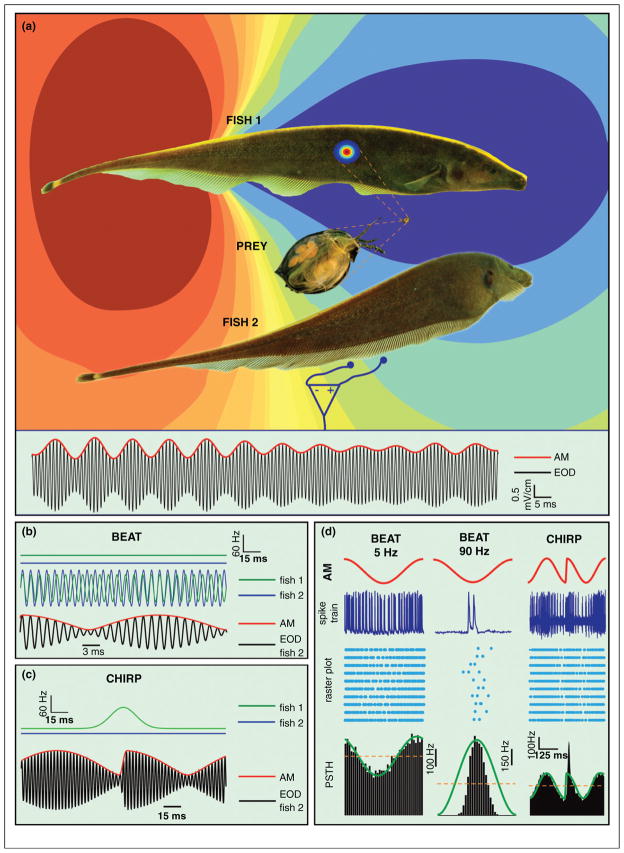
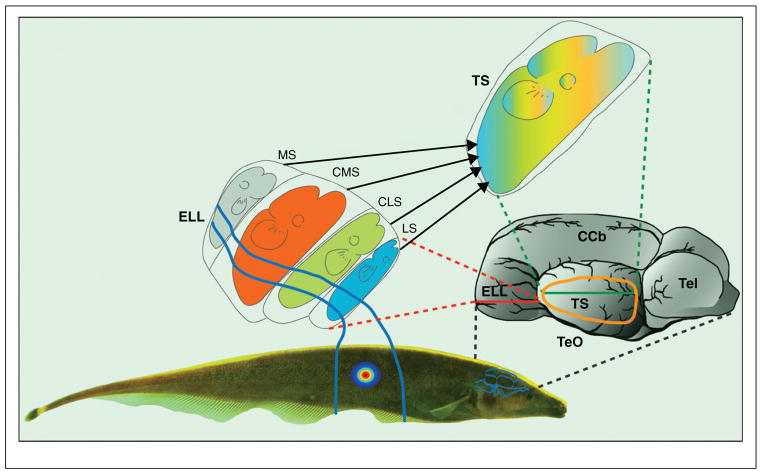
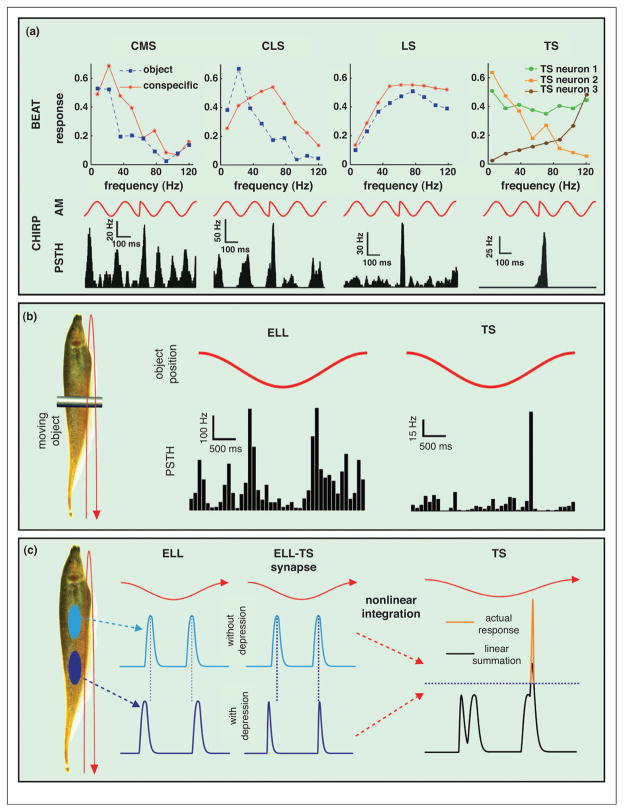
References
-
- Barlow HB. Single Units and sensation: a neuron doctrine for perceptual psychology. Perception. 1972;1:371–394. - PubMed
-
- Rolls ET, Tovee MJ. Sparseness of the neuronal representation of stimuli in the primate temporal visual cortex. J Neurophysiol. 1995;73:713–726. - PubMed
-
- Quiroga RQ, Reddy L, Kreiman G, Koch C, Fried I. Invariant visual representation by single neurons in the human brain [see comment] Nature. 2005;435:1102–1107. - PubMed
-
- Perez-Orive J, Mazor O, Turner GC, Cassenaer S, Wilson RI, Laurent G. Oscillations and sparsening of odor representations in the mushroom body. Science. 2002;297:359–365. - PubMed
-
- Olshausen BA, Field DJ. Sparse coding of sensory inputs. Curr Opin Neurobiol. 2004;14:481–487. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
