

Spikes alone do not behavior make: why neuroscience needs biomechanics
- PMID: 21683575
- PMCID: PMC3183174
- DOI: 10.1016/j.conb.2011.05.017
Spikes alone do not behavior make: why neuroscience needs biomechanics
Abstract
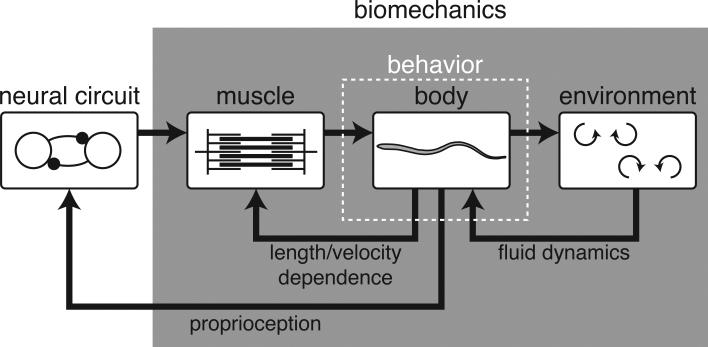
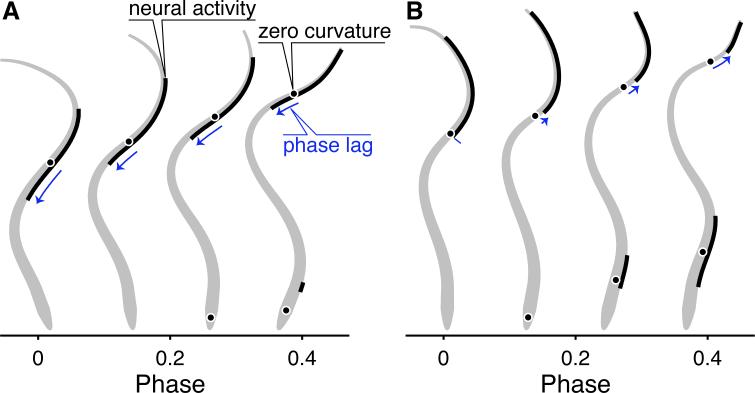
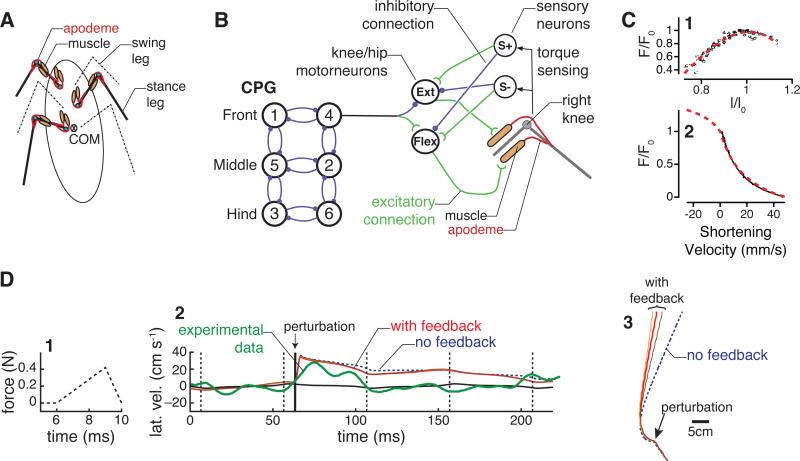
Neural circuits do not function in isolation; they interact with the physical world, accepting sensory inputs and producing outputs via muscles. Since both these pathways are constrained by physics, the activity of neural circuits can only be understood by considering biomechanics of muscles, bodies, and the exterior world. We discuss how animal bodies have natural stable motions that require relatively little activation or control from the nervous system. The nervous system can substantially alter these motions, by subtly changing mechanical properties such as body or leg stiffness. Mechanics can also provide robustness to perturbations without sensory reflexes. By considering a complete neuromechanical system, neuroscientists and biomechanicians together can provide a more integrated view of neural circuitry and behavior.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Domenici P, Blake RW. The kinematics and performance of fish fast-start swimming. J Exp Biol. 1997;200:1165–1178. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
