

A Smad action turnover switch operated by WW domain readers of a phosphoserine code
- PMID: 21685363
- PMCID: PMC3127429
- DOI: 10.1101/gad.2060811
A Smad action turnover switch operated by WW domain readers of a phosphoserine code
Abstract
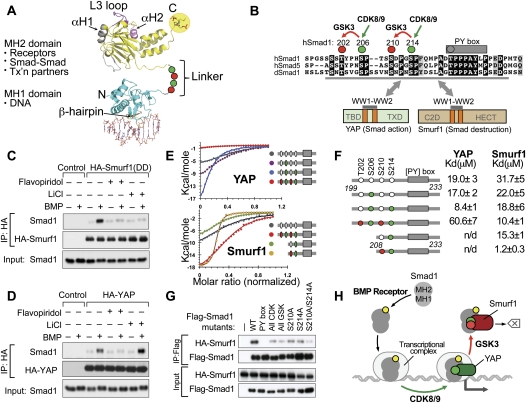
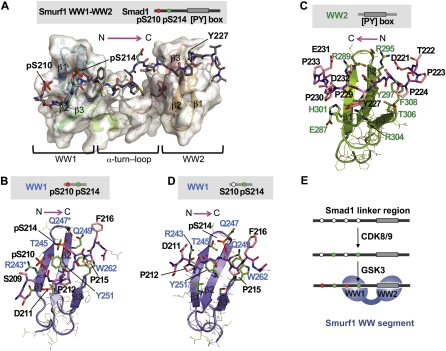
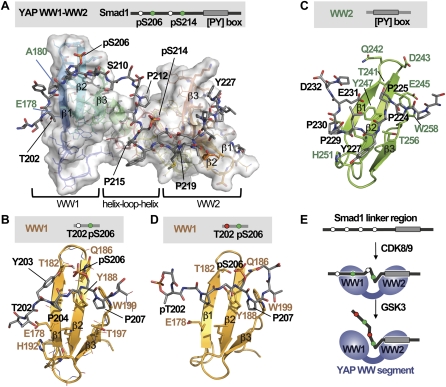
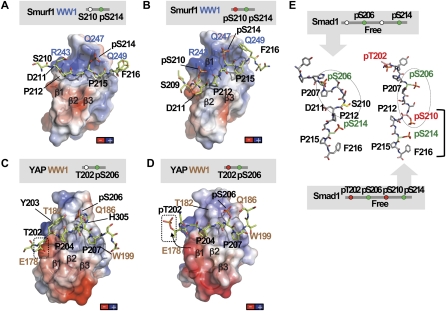
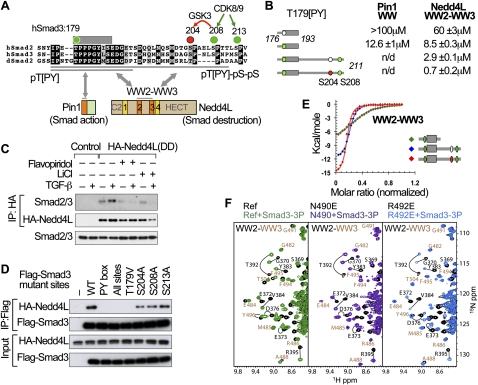
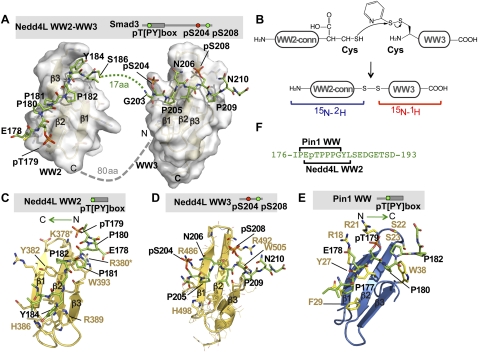
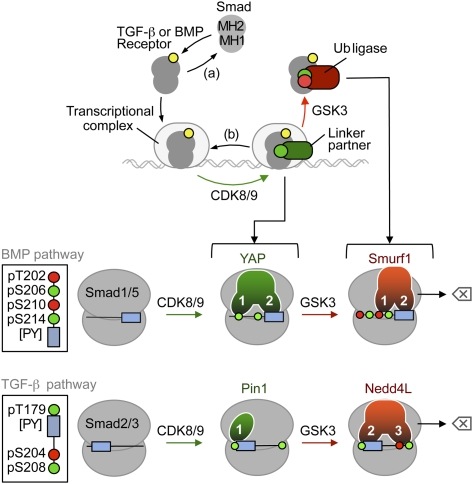
When directed to the nucleus by TGF-β or BMP signals, Smad proteins undergo cyclin-dependent kinase 8/9 (CDK8/9) and glycogen synthase kinase-3 (GSK3) phosphorylations that mediate the binding of YAP and Pin1 for transcriptional action, and of ubiquitin ligases Smurf1 and Nedd4L for Smad destruction. Here we demonstrate that there is an order of events-Smad activation first and destruction later-and that it is controlled by a switch in the recognition of Smad phosphoserines by WW domains in their binding partners. In the BMP pathway, Smad1 phosphorylation by CDK8/9 creates binding sites for the WW domains of YAP, and subsequent phosphorylation by GSK3 switches off YAP binding and adds binding sites for Smurf1 WW domains. Similarly, in the TGF-β pathway, Smad3 phosphorylation by CDK8/9 creates binding sites for Pin1 and GSK3, then adds sites to enhance Nedd4L binding. Thus, a Smad phosphoserine code and a set of WW domain code readers provide an efficient solution to the problem of coupling TGF-β signal delivery to turnover of the Smad signal transducers.
Figures
References
-
- Baca M, Muir TW, Schnölzer M, Kent SBH 1995. Chemical ligation of cysteine- containing peptides: synthesis of a 22 kDa tethered dimer of HIV-1 protease. J Am Chem Soc 116: 10797–10798
-
- Bartels C, Xia T-H, Billeter M, Güntert P, Wüthrich K 1995. The program XEASY for computer-supported NMR spectral analysis of biological macromolecules. J Biol NMR 5: 1–10 - PubMed
-
- Brünger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, et al. 1998. Crystallography and NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr 54: 905–921 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous