

Analysis of protein dynamics at active, stalled, and collapsed replication forks
- PMID: 21685366
- PMCID: PMC3127432
- DOI: 10.1101/gad.2053211
Analysis of protein dynamics at active, stalled, and collapsed replication forks
Abstract
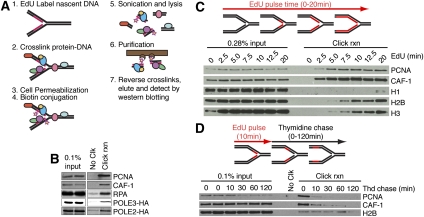
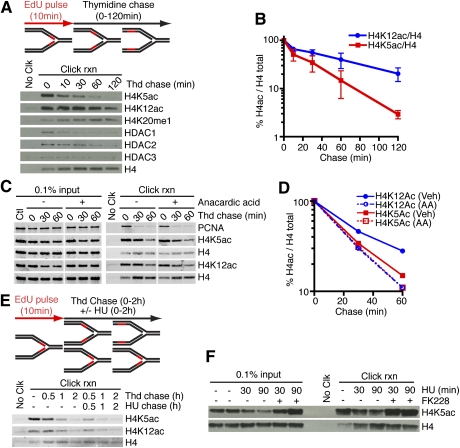
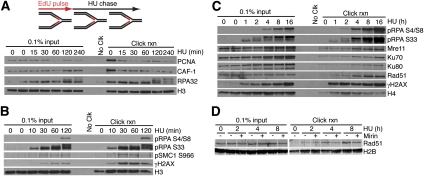
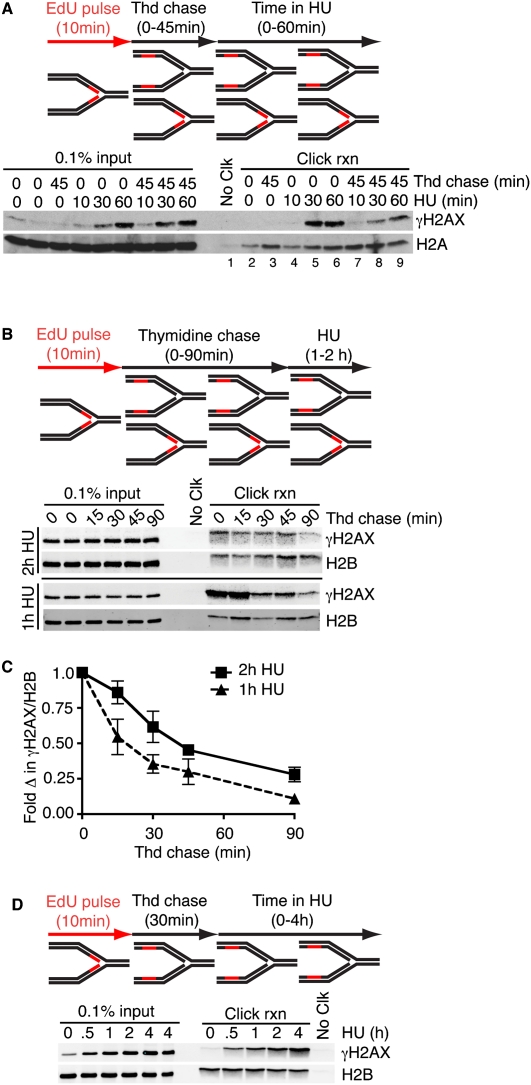
Successful DNA replication and packaging of newly synthesized DNA into chromatin are essential to maintain genome integrity. Defects in the DNA template challenge genetic and epigenetic inheritance. Unfortunately, tracking DNA damage responses (DDRs), histone deposition, and chromatin maturation at replication forks is difficult in mammalian cells. Here we describe a technology called iPOND (isolation of proteins on nascent DNA) to analyze proteins at active and damaged replication forks at high resolution. Using this methodology, we define the timing of histone deposition and chromatin maturation. Class 1 histone deacetylases are enriched at replisomes and remove predeposition marks on histone H4. Chromatin maturation continues even when decoupled from replisome movement. Furthermore, fork stalling causes changes in the recruitment and phosphorylation of proteins at the damaged fork. Checkpoint kinases catalyze H2AX phosphorylation, which spreads from the stalled fork to include a large chromatin domain even prior to fork collapse and double-strand break formation. Finally, we demonstrate a switch in the DDR at persistently stalled forks that includes MRE11-dependent RAD51 assembly. These data reveal a dynamic recruitment of proteins and post-translational modifications at damaged forks and surrounding chromatin. Furthermore, our studies establish iPOND as a useful methodology to study DNA replication and chromatin maturation.
Figures
References
-
- Ahmad A, Takami Y, Nakayama T 1999. WD repeats of the p48 subunit of chicken chromatin assembly factor-1 required for in vitro interaction with chicken histone deacetylase-2. J Biol Chem 274: 16646–16653 - PubMed
-
- Anantha RW, Vassin VM, Borowiec JA 2007. Sequential and synergistic modification of human RPA stimulates chromosomal DNA repair. J Biol Chem 282: 35910–35923 - PubMed
-
- Berkovich E, Monnat RJ Jr, Kastan MB 2007. Roles of ATM and NBS1 in chromatin structure modulation and DNA double-strand break repair. Nat Cell Biol 9: 683–690 - PubMed
-
- Berkovich E, Monnat RJ Jr, Kastan MB 2008. Assessment of protein dynamics and DNA repair following generation of DNA double-strand breaks at defined genomic sites. Nat Protoc 3: 915–922 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials