

Imaging analysis of clock neurons reveals light buffers the wake-promoting effect of dopamine
- PMID: 21685918
- PMCID: PMC3424274
- DOI: 10.1038/nn.2860
Imaging analysis of clock neurons reveals light buffers the wake-promoting effect of dopamine
Abstract
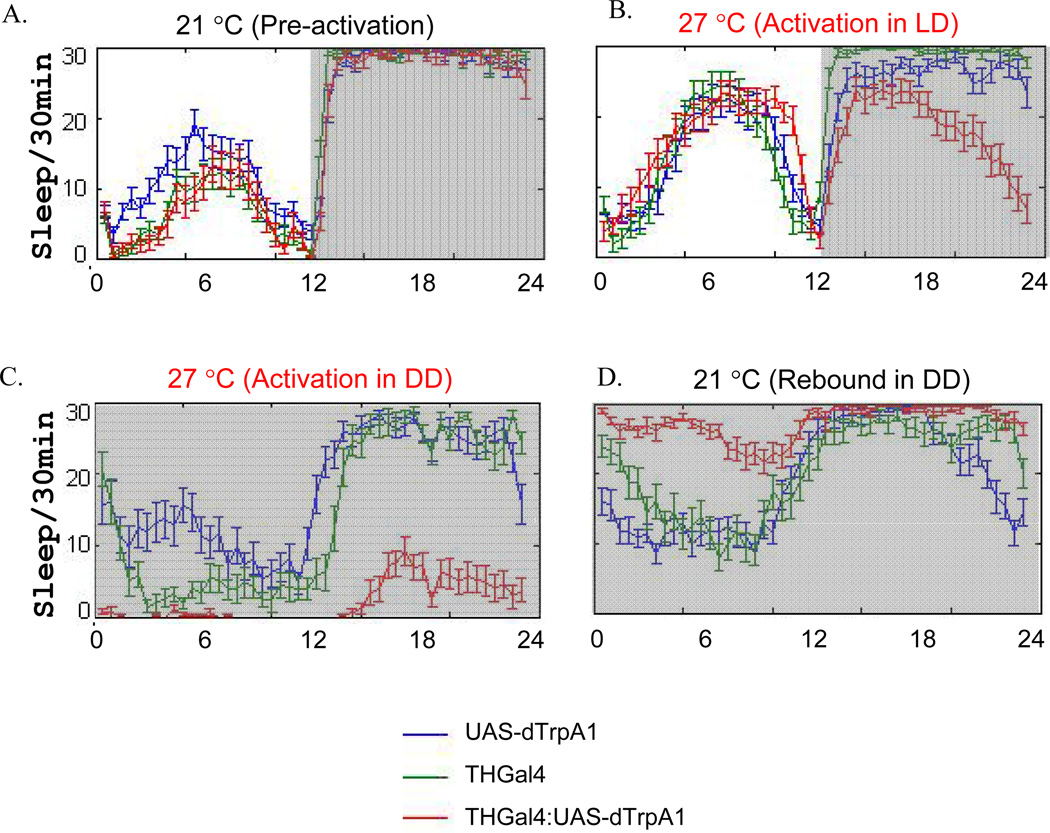
How animals maintain proper amounts of sleep yet remain flexible to changes in environmental conditions remains unknown. We found that environmental light suppressed the wake-promoting effects of dopamine in fly brains. The ten large lateral-ventral neurons (l-LNvs), a subset of clock neurons, are wake-promoting and respond to dopamine, octopamine and light. Behavioral and imaging analyses suggested that dopamine is a stronger arousal signal than octopamine. Notably, light exposure not only suppressed l-LNv responses, but also synchronized responses of neighboring l-LNvs. This regulation occurred by distinct mechanisms: light-mediated suppression of octopamine responses was regulated by the circadian clock, whereas light regulation of dopamine responses occurred by upregulation of inhibitory dopamine receptors. Plasticity therefore alters the relative importance of diverse cues on the basis of the environmental mix of stimuli. The regulatory mechanisms described here may contribute to the control of sleep stability while still allowing behavioral flexibility.
Figures





References
-
- Ganguly-Fitzgerald I, Donlea J, Shaw PJ. Waking experience affects sleep need in Drosophila. Science. 2006;313:1775–1781. - PubMed
-
- Ho KS, Sehgal A. Drosophila melanogaster: an insect model for fundamental studies of sleep. Methods Enzymol. 2005;393:772–793. - PubMed
-
- Andretic R, van Swinderen B, Greenspan RJ. Dopaminergic modulation of arousal in Drosophila. Curr Biol. 2005;15:1165–1175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
