

Trypanosoma cruzi Coexpressing Ornithine Decarboxylase and Green Fluorescence Proteins as a Tool to Study the Role of Polyamines in Chagas Disease Pathology
- PMID: 21687606
- PMCID: PMC3112526
- DOI: 10.4061/2011/657460
Trypanosoma cruzi Coexpressing Ornithine Decarboxylase and Green Fluorescence Proteins as a Tool to Study the Role of Polyamines in Chagas Disease Pathology
Abstract
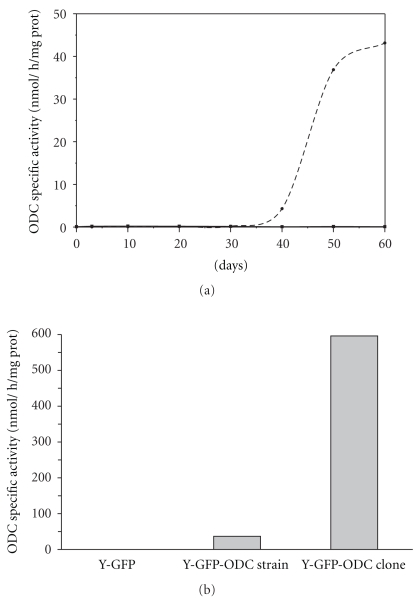
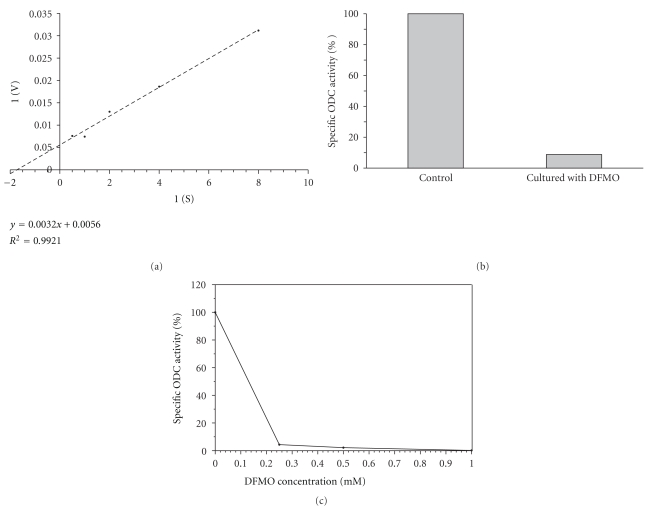
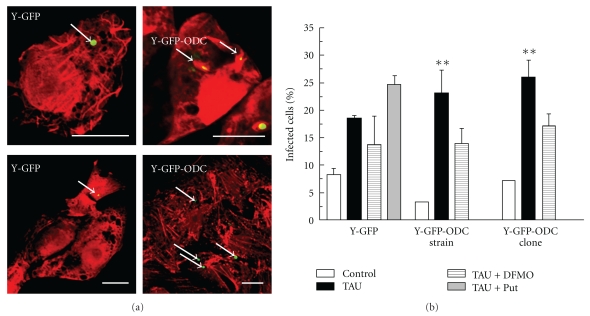
Polyamines are essential for Trypanosoma cruzi, the causative agent of Chagas disease. As T. cruzi behaves as a natural auxotrophic organism, it relies on host polyamines biosynthesis. In this paper we obtained a double-transfected T. cruzi parasite that expresses the green fluorescent protein (GFP) and a heterologous ornithine decarboxylase (ODC), used itself as a novel selectable marker. These autotrophic and fluorescent parasites were characterized; the ODC presented an apparent Km for ornithine of 0.51 ± 0.16 mM and an estimated V(max) value of 476.2 nmoles/h/mg of protein. These expressing ODC parasites showed higher metacyclogenesis capacity than the auxotrophic counterpart, supporting the idea that polyamines are engaged in this process. This double-transfected T. cruzi parasite results in a powerful tool-easy to follow by its fluorescence-to study the role of polyamines in Chagas disease pathology and in related processes such as parasite survival, invasion, proliferation, metacyclogenesis, and tissue spreading.
Figures
References
-
- Heby O, Persson L, Rentala M. Targeting the polyamine biosynthetic enzymes: a promising approach to therapy of African sleeping sickness, Chagas’ disease, and leishmaniasis. Amino Acids. 2007;33(2):359–366. - PubMed
-
- Schofield CJ, Maudlin I. Trypanosomiasis control. International Journal for Parasitology. 2001;31(5-6):615–620. - PubMed
-
- Andrade LO, Andrews NW. Opinion: the Trypanosoma cruzi—host-cell interplay: location, invasion, retention. Nature Reviews Microbiology. 2005;3(10):819–823. - PubMed
-
- Tyler KM, Engman DM. The life cycle of Trypanosoma cruzi revisited. International Journal for Parasitology. 2001;31(5-6):472–481. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
