

Prokineticin 2 regulates the electrical activity of rat suprachiasmatic nuclei neurons
- PMID: 21687716
- PMCID: PMC3110640
- DOI: 10.1371/journal.pone.0020263
Prokineticin 2 regulates the electrical activity of rat suprachiasmatic nuclei neurons
Abstract
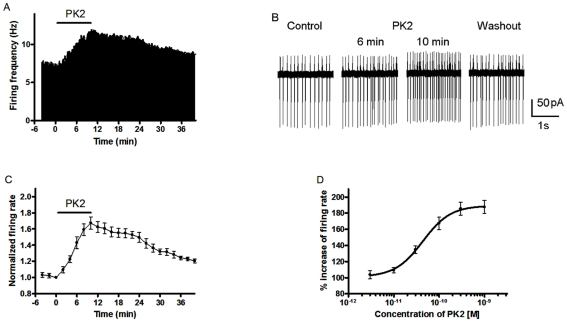
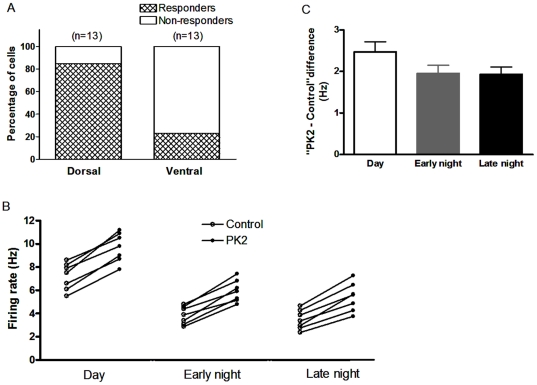
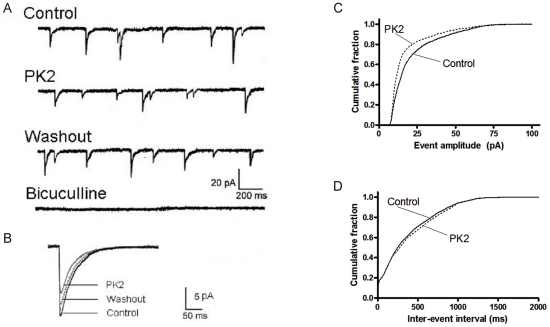
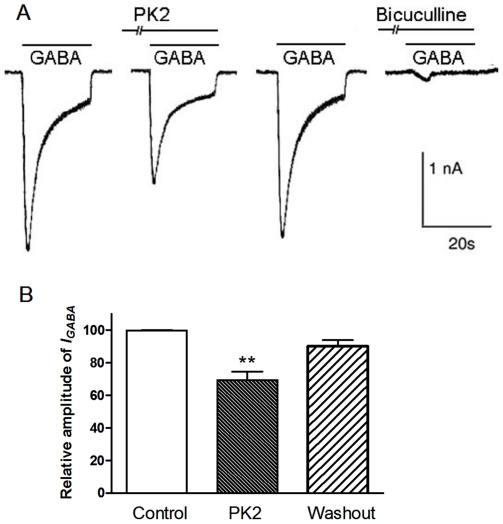
Neuropeptide signaling plays roles in coordinating cellular activities and maintaining robust oscillations within the mammalian suprachiasmatic nucleus (SCN). Prokineticin2 (PK2) is a signaling molecule from the SCN and involves in the generation of circadian locomotor activity. Prokineticin receptor 2 (PKR2), a receptor for PK2, has been shown to be expressed in the SCN. However, very little is known about the cellular action of PK2 within the SCN. In the present study, we investigated the effect of PK2 on spontaneous firing and miniature inhibitory postsynaptic currents (mIPSCs) using whole cell patch-clamp recording in the SCN slices. PK2 dose-dependently increased spontaneous firing rates in most neurons from the dorsal SCN. PK2 acted postsynaptically to reduce γ-aminobutyric acid (GABA)-ergic function within the SCN, and PK2 reduced the amplitude but not frequency of mIPSCs. Furthermore, PK2 also suppressed exogenous GABA-induced currents. And the inhibitory effect of PK2 required PKC activation in the postsynaptic cells. Our data suggest that PK2 could alter cellular activities within the SCN and may influence behavioral and physiological rhythms.
Conflict of interest statement
Figures
Similar articles
-
Distinct localization of prokineticin 2 and prokineticin receptor 2 mRNAs in the rat suprachiasmatic nucleus.Eur J Neurosci. 2006 Jun;23(11):2959-70. doi: 10.1111/j.1460-9568.2006.04834.x. Eur J Neurosci. 2006. PMID: 16819985
-
Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus.Nature. 2002 May 23;417(6887):405-10. doi: 10.1038/417405a. Nature. 2002. PMID: 12024206
-
Prokineticin 2 protein is diurnally expressed in PER2-containing clock neurons in the mouse suprachiasmatic nucleus.Peptides. 2025 Jan;183:171339. doi: 10.1016/j.peptides.2024.171339. Epub 2025 Jan 2. Peptides. 2025. PMID: 39755259
-
The circadian output signals from the suprachiasmatic nuclei.Prog Brain Res. 2012;199:119-127. doi: 10.1016/B978-0-444-59427-3.00028-9. Prog Brain Res. 2012. PMID: 22877662 Review.
-
Prokineticin 2 and circadian clock output.FEBS J. 2005 Nov;272(22):5703-9. doi: 10.1111/j.1742-4658.2005.04984.x. FEBS J. 2005. PMID: 16279936 Free PMC article. Review.
Cited by
-
Functional rescue of Kallmann syndrome-associated prokineticin receptor 2 (PKR2) mutants deficient in trafficking.J Biol Chem. 2014 May 30;289(22):15518-26. doi: 10.1074/jbc.M114.556381. Epub 2014 Apr 21. J Biol Chem. 2014. PMID: 24753254 Free PMC article.
-
Temporal variations in presynaptic release probability in the lateral habenula.Sci Rep. 2017 Jan 20;7:40866. doi: 10.1038/srep40866. Sci Rep. 2017. PMID: 28106159 Free PMC article.
-
Ion Channels Controlling Circadian Rhythms in Suprachiasmatic Nucleus Excitability.Physiol Rev. 2020 Oct 1;100(4):1415-1454. doi: 10.1152/physrev.00027.2019. Epub 2020 Mar 12. Physiol Rev. 2020. PMID: 32163720 Free PMC article. Review.
-
Signaling role of prokineticin 2 on the estrous cycle of female mice.PLoS One. 2014 Mar 14;9(3):e90860. doi: 10.1371/journal.pone.0090860. eCollection 2014. PLoS One. 2014. PMID: 24633064 Free PMC article.
-
Controlling the activation of the Bv8/prokineticin system reduces neuroinflammation and abolishes thermal and tactile hyperalgesia in neuropathic animals.Br J Pharmacol. 2014 Nov;171(21):4850-65. doi: 10.1111/bph.12793. Epub 2014 Sep 5. Br J Pharmacol. 2014. PMID: 24902717 Free PMC article.
References
-
- Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002;418:935–941. - PubMed
-
- Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock. Hum Mol Genet. 2006;15 Spec No 2:R271–277. - PubMed
-
- Kuhlman SJ, McMahon DG. Encoding the ins and outs of circadian pacemaking. J Biol Rhythms. 2006;21:470–481. - PubMed
-
- Morin LP, Allen CN. The circadian visual system, 2005. Brain Res Rev. 2006;51:1–60. - PubMed
-
- Brown TM, Piggins HD. Electrophysiology of the suprachiasmatic circadian clock. Prog Neurobiol. 2007;82:229–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
