

Evolutionary diversity and developmental regulation of X-chromosome inactivation
- PMID: 21687993
- PMCID: PMC3132430
- DOI: 10.1007/s00439-011-1029-2
Evolutionary diversity and developmental regulation of X-chromosome inactivation
Abstract
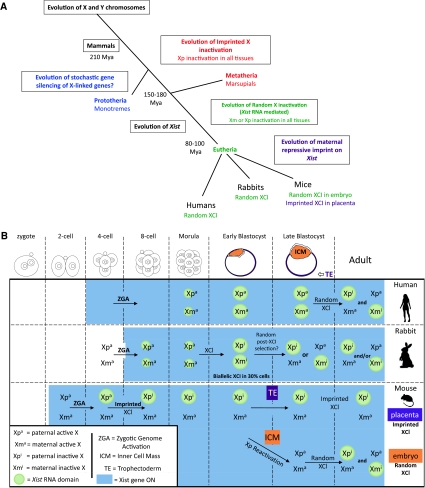
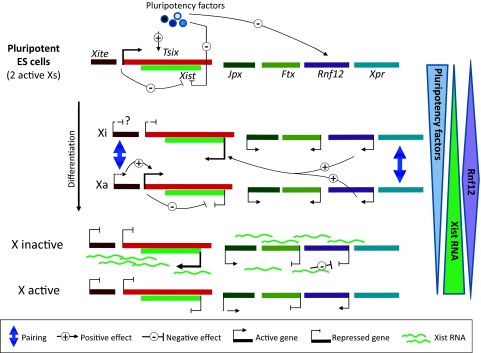
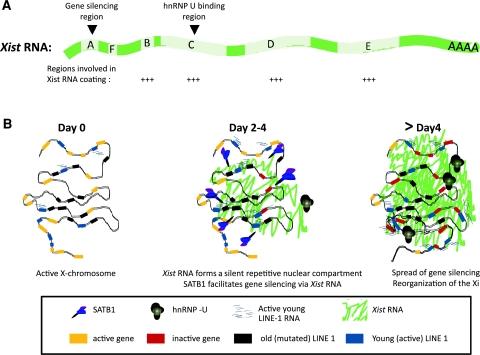
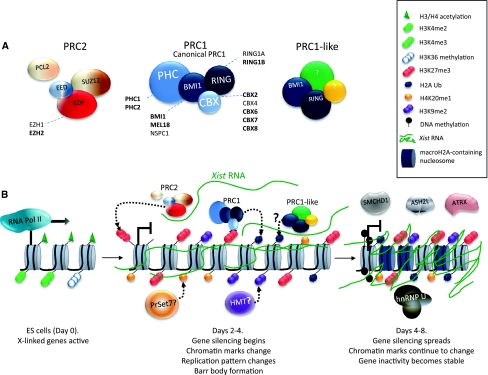
X-chromosome inactivation (XCI) results in the transcriptional silencing of one X-chromosome in females to attain gene dosage parity between XX female and XY male mammals. Mammals appear to have developed rather diverse strategies to initiate XCI in early development. In placental mammals XCI depends on the regulatory noncoding RNA X-inactive specific transcript (Xist), which is absent in marsupials and monotremes. Surprisingly, even placental mammals show differences in the initiation of XCI in terms of Xist regulation and the timing to acquire dosage compensation. Despite this, all placental mammals achieve chromosome-wide gene silencing at some point in development, and this is maintained by epigenetic marks such as chromatin modifications and DNA methylation. In this review, we will summarise recent findings concerning the events that occur downstream of Xist RNA coating of the inactive X-chromosome (Xi) to ensure its heterochromatinization and the maintenance of the inactive state in the mouse and highlight similarities and differences between mammals.
Figures
References
-
- Augui S, Filion GJ, Huart S, Nora E, Guggiari M, Maresca M, Stewart AF, Heard E. Sensing X chromosome pairs before X inactivation via a novel X-pairing region of the Xic. Science. 2007;318:1632–1636. - PubMed
-
- Bacher CP, Guggiari M, Brors B, Augui S, Clerc P, Avner P, Eils R, Heard E. Transient colocalization of X-inactivation centres accompanies the initiation of X inactivation. Nat Cell Biol. 2006;8:293–299. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
