

Prestin-driven cochlear amplification is not limited by the outer hair cell membrane time constant
- PMID: 21689600
- PMCID: PMC3143834
- DOI: 10.1016/j.neuron.2011.04.024
Prestin-driven cochlear amplification is not limited by the outer hair cell membrane time constant
Abstract
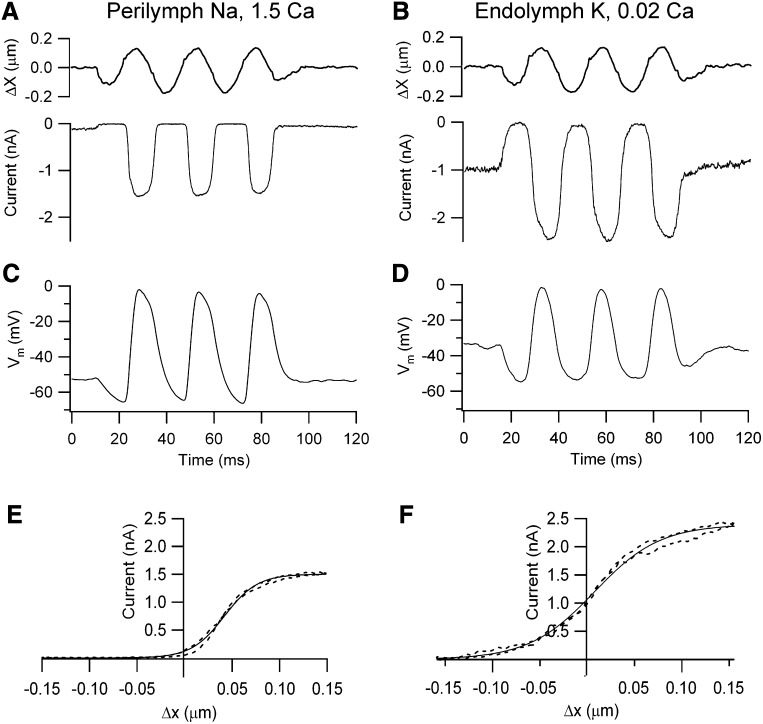
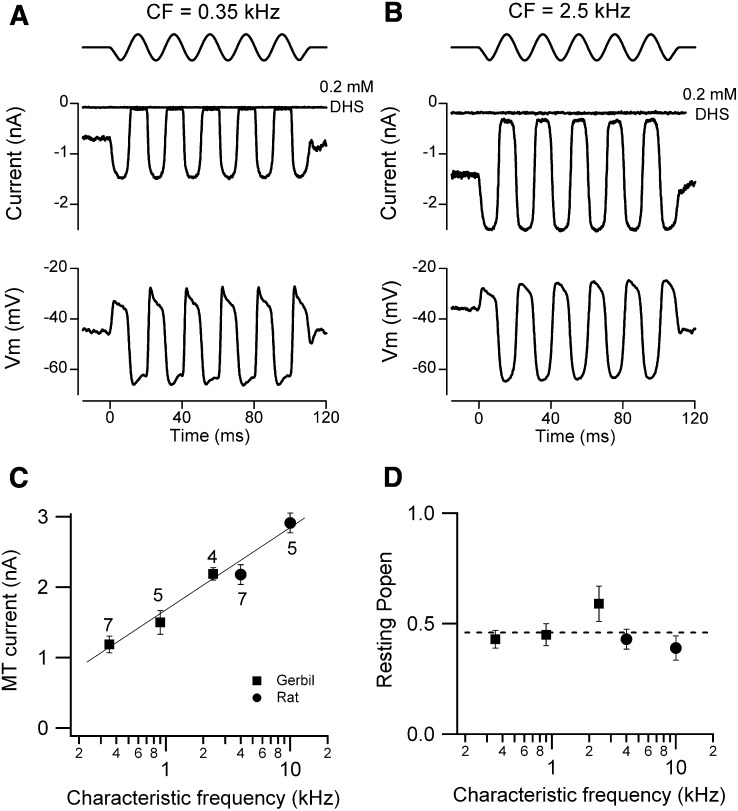
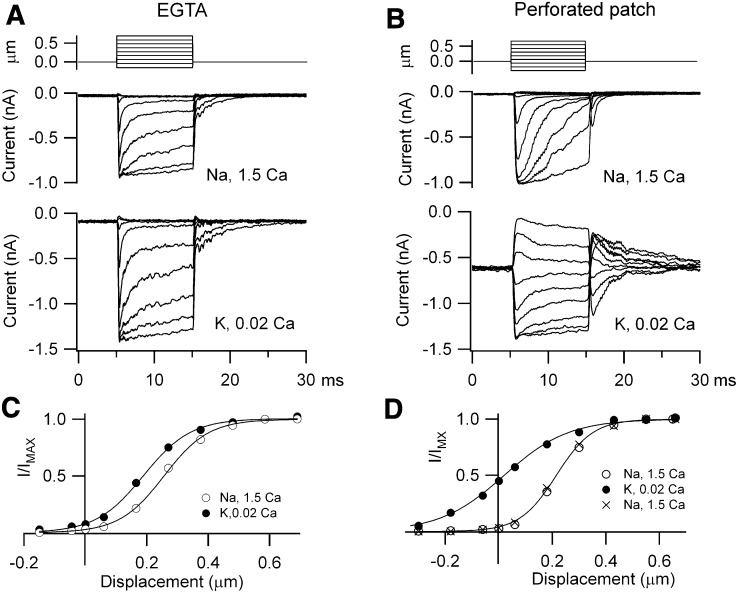
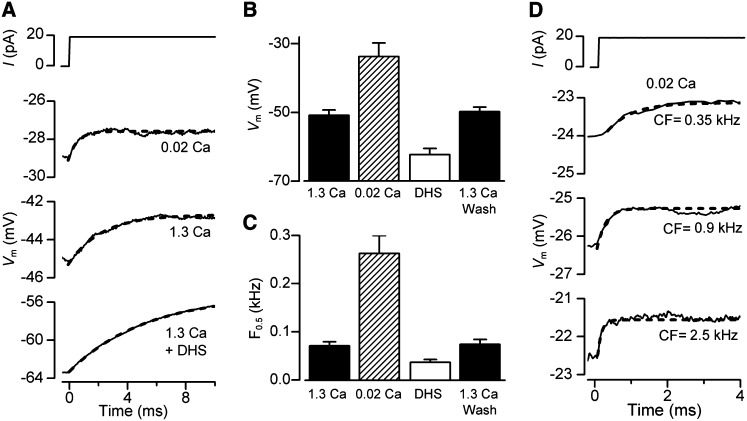
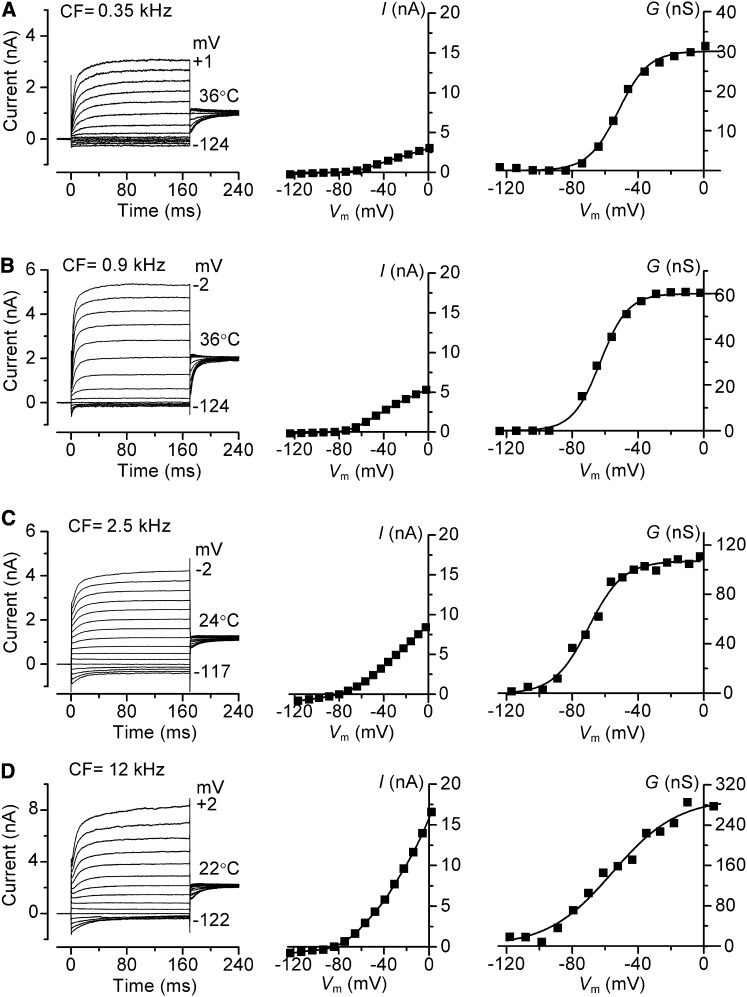
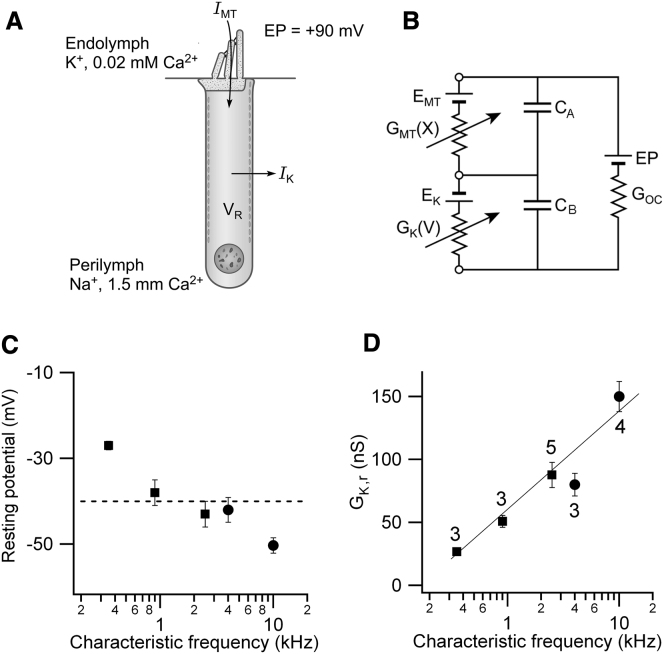
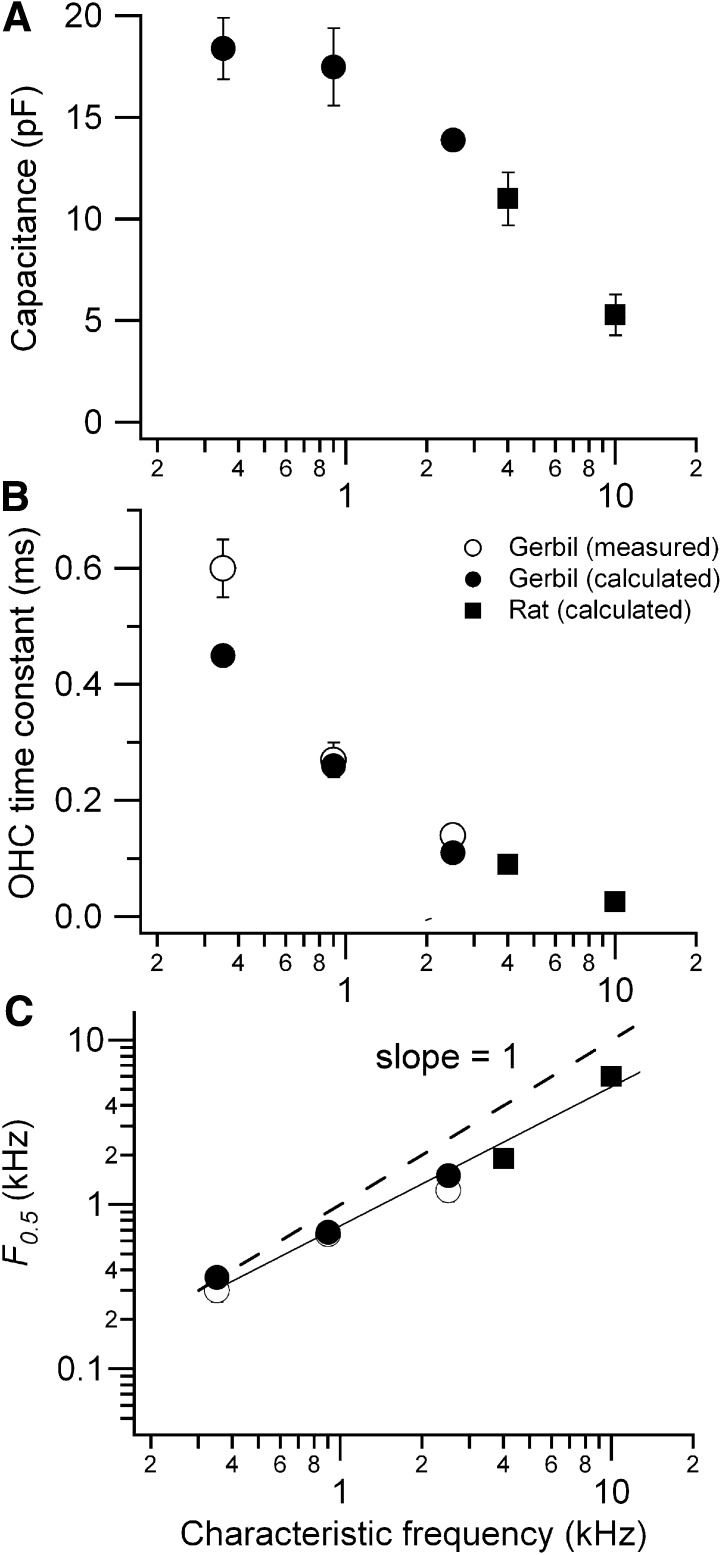
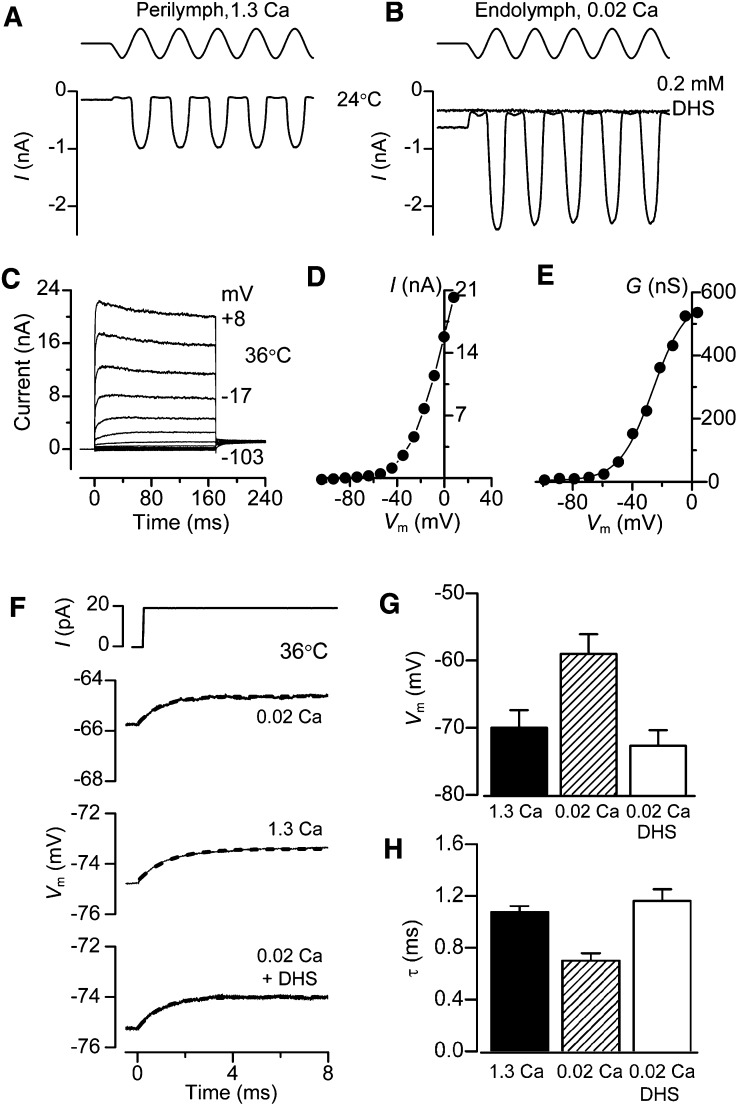
Outer hair cells (OHCs) provide amplification in the mammalian cochlea using somatic force generation underpinned by voltage-dependent conformational changes of the motor protein prestin. However, prestin must be gated by changes in membrane potential on a cycle-by-cycle basis and the periodic component of the receptor potential may be greatly attenuated by low-pass filtering due to the OHC time constant (τ(m)), questioning the functional relevance of this mechanism. Here, we measured τ(m) from OHCs with a range of characteristic frequencies (CF) and found that, at physiological endolymphatic calcium concentrations, approximately half of the mechanotransducer (MT) channels are opened at rest, depolarizing the membrane potential to near -40 mV. The depolarized resting potential activates a voltage-dependent K+ conductance, thus minimizing τ(m) and expanding the membrane filter so there is little receptor potential attenuation at the cell's CF. These data suggest that minimal τ(m) filtering in vivo ensures optimal activation of prestin.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Pushing the envelope of sound.Neuron. 2011 Jun 23;70(6):1021-2. doi: 10.1016/j.neuron.2011.06.002. Neuron. 2011. PMID: 21689589
References
-
- Ashmore J. Cochlear outer hair cell motility. Physiol. Rev. 2008;88:173–210. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
