

Notch: architect, landscaper, and guardian of the intestine
- PMID: 21689653
- PMCID: PMC4050496
- DOI: 10.1053/j.gastro.2011.06.003
Notch: architect, landscaper, and guardian of the intestine
Abstract
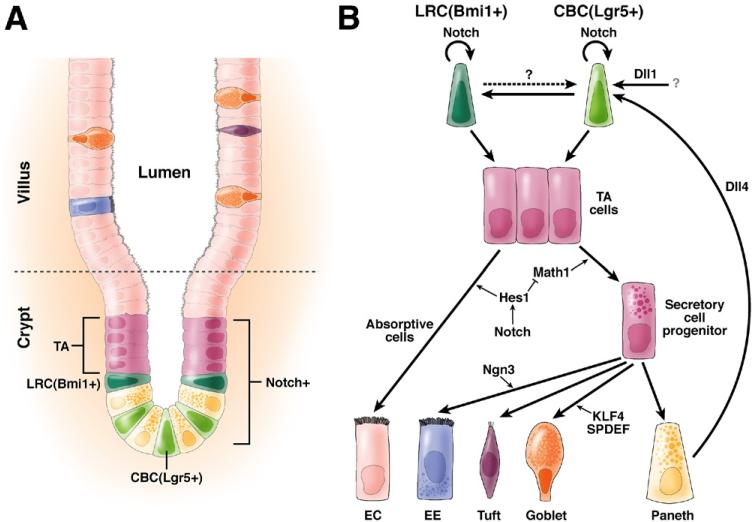
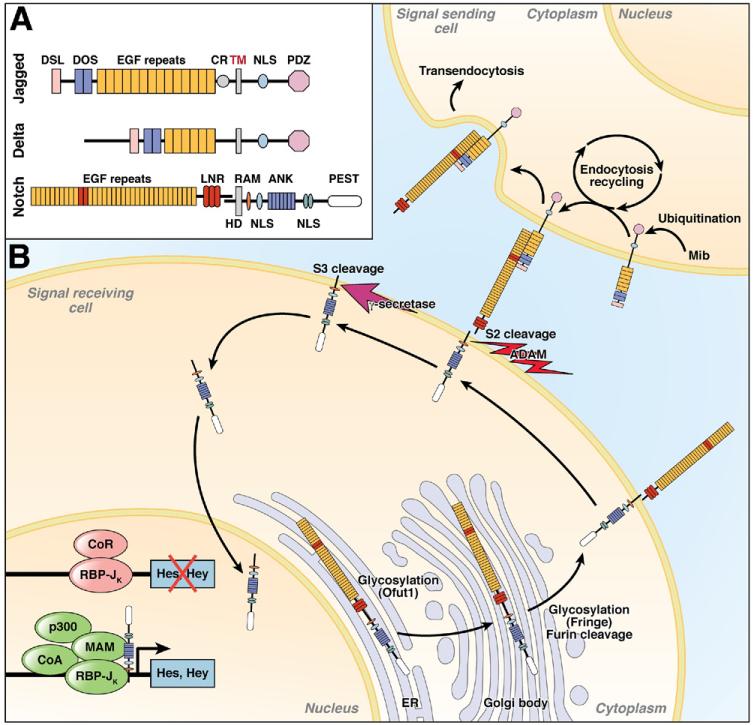
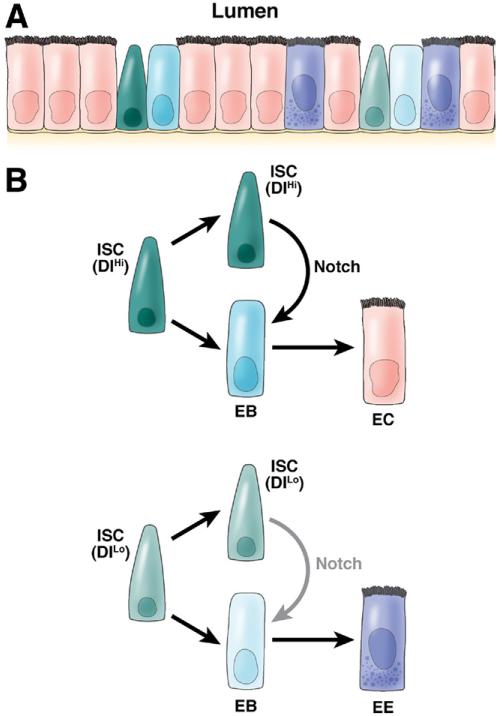
In the past decade, enormous progress has been made in understanding the role of stem cells in physiologic tissue renewal and in pathologic processes such as cancer. These findings have shed light on the identity and biological properties of such cells and the intrinsic and extrinsic signals that balance stem cell self-renewal with differentiation. With its astonishing self-renewal capacity, the intestinal epithelium has provided a unique model to study stem cell biology, lineage specification, and cancer. Here we review the role of Notch signaling in physiologic cell renewal and differentiation in the intestine as well as during its malignant transformation.
Copyright © 2011 AGA Institute. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Sancho E, Batlle E, Clevers H. Signaling pathways in intestinal development and cancer. Annu Rev Cell Dev Biol. 2004;20:695–723. - PubMed
-
- Cheng H, Bjerknes M. Cell production in mouse intestinal epithelium measured by stathmokinetic flow cytometry and Coulter particle counting. Anat Rec. 1983;207:427–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
