

Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis
- PMID: 21690308
- PMCID: PMC3216325
- DOI: 10.1083/jcb.201103076
Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis
Abstract
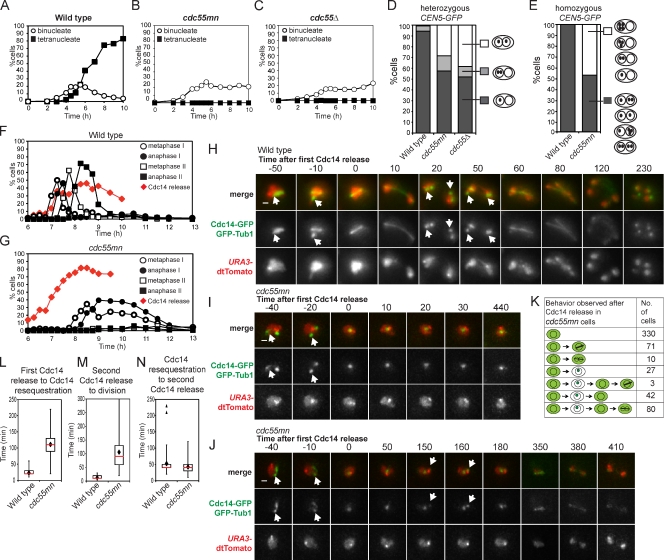
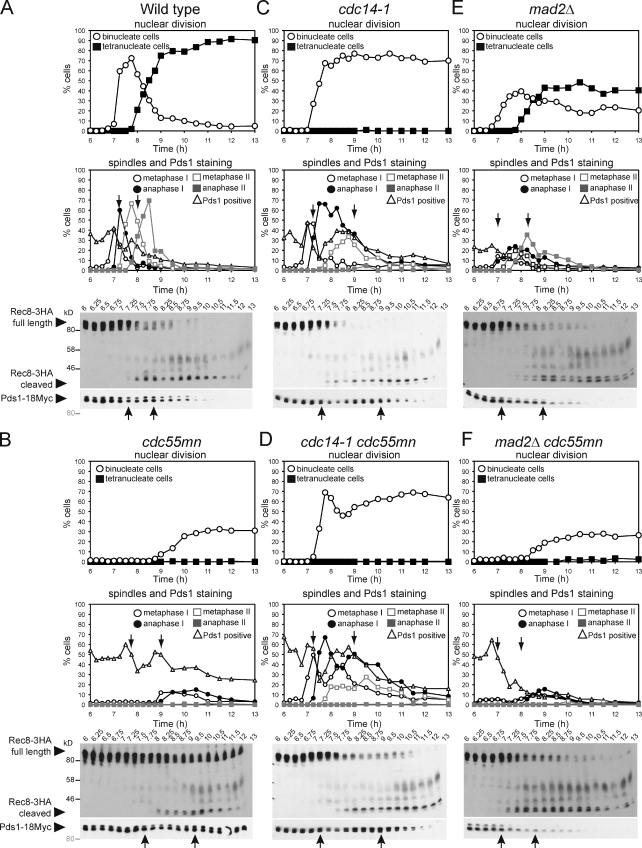
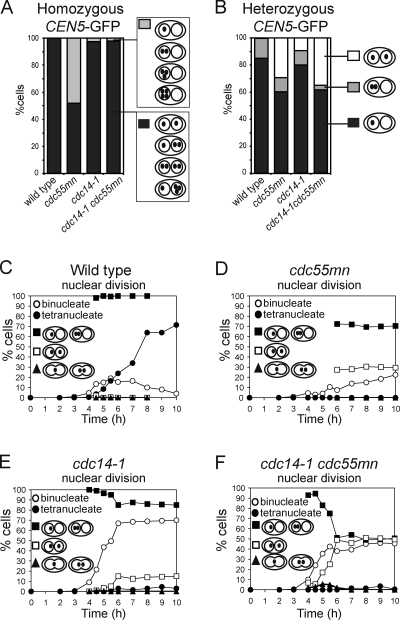
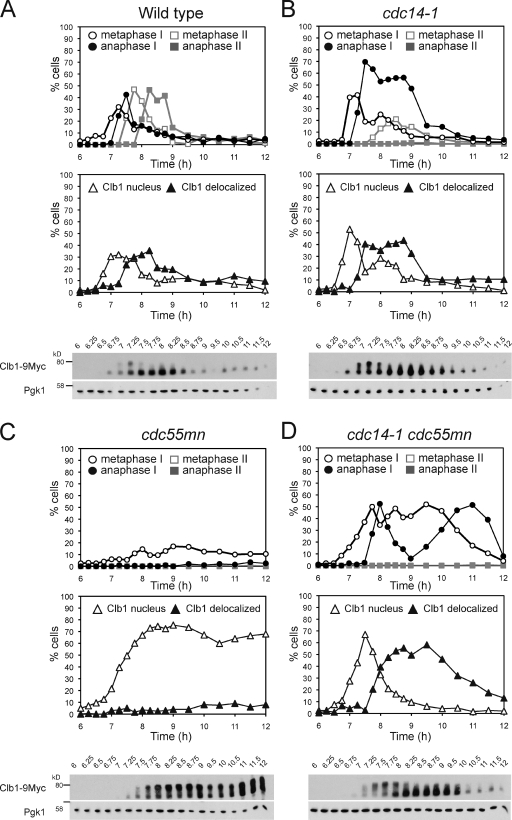
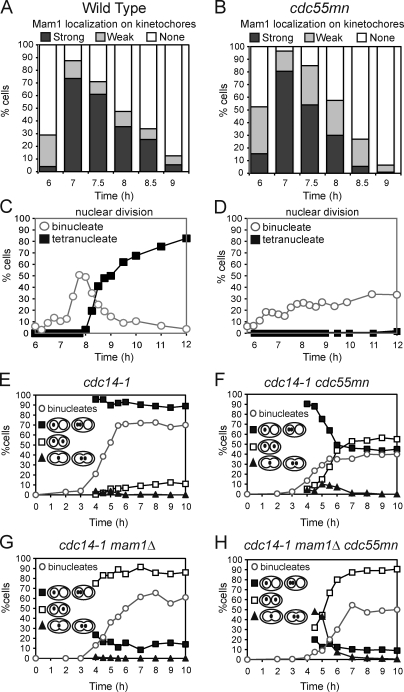
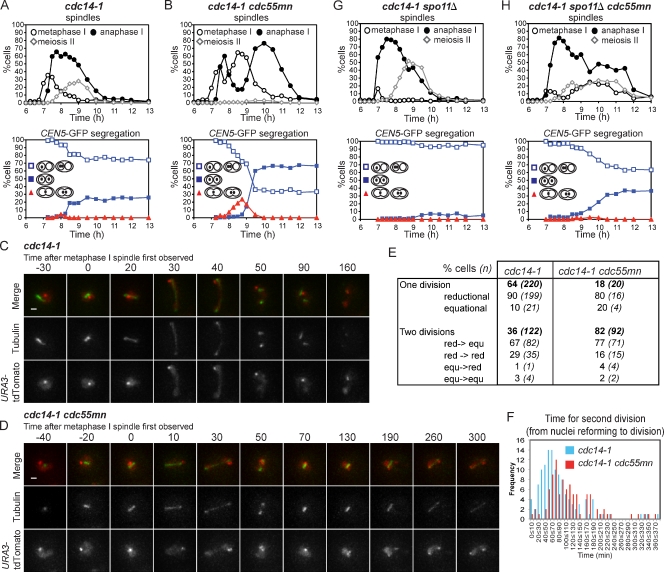
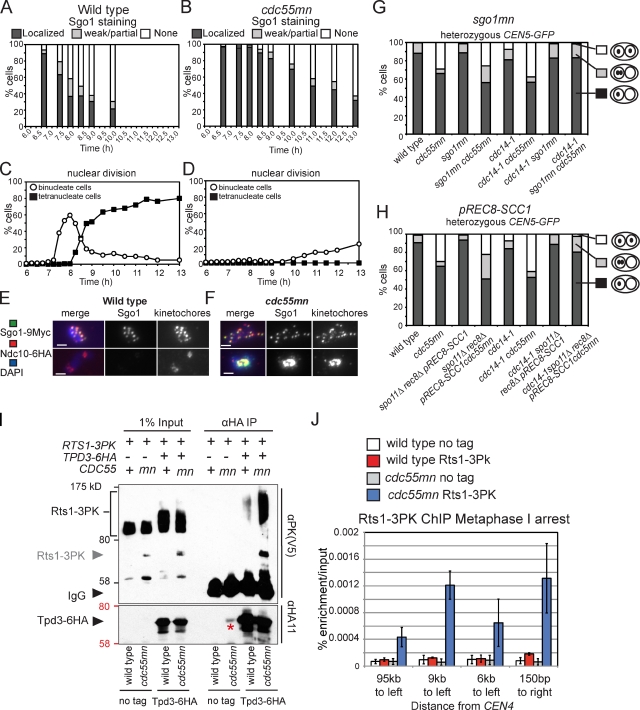
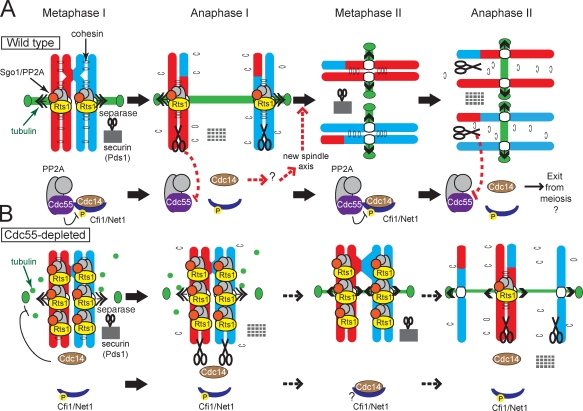
During meiosis, two consecutive nuclear divisions follow a single round of deoxyribonucleic acid replication. In meiosis I, homologues are segregated, whereas in meiosis II, sister chromatids are segregated. This requires that the sequential assembly and dissolution of specialized chromosomal factors are coordinated with two rounds of spindle assembly and disassembly. How these events are coupled is unknown. In this paper, we show, in budding yeast, that the protein phosphatase 2A regulatory subunit Cdc55 couples the loss of linkages between chromosomes with nuclear division by restraining two other phosphatases, Cdc14 and PP2A(Rts1). Cdc55 maintains Cdc14 sequestration in the nucleolus during early meiosis, and this is essential for the assembly of the meiosis I spindle but not for chromosomes to separate. Cdc55 also limits the formation of PP2A holocomplexes containing the alternative regulatory subunit Rts1, which is crucial for the timely dissolution of sister chromatid cohesion. Therefore, Cdc55 orders passage through the meiotic divisions by ensuring a balance of phosphatases.
Figures
References
-
- Buonomo S.B., Rabitsch K.P., Fuchs J., Gruber S., Sullivan M., Uhlmann F., Petronczki M., Tóth A., Nasmyth K. 2003. Division of the nucleolus and its release of CDC14 during anaphase of meiosis I depends on separase, SPO12, and SLK19. Dev. Cell. 4:727–739 10.1016/S1534-5807(03)00129-1 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
