

How to make stripes: deciphering the transition from non-periodic to periodic patterns in Drosophila segmentation
- PMID: 21693522
- PMCID: PMC3119311
- DOI: 10.1242/dev.062141
How to make stripes: deciphering the transition from non-periodic to periodic patterns in Drosophila segmentation
Abstract
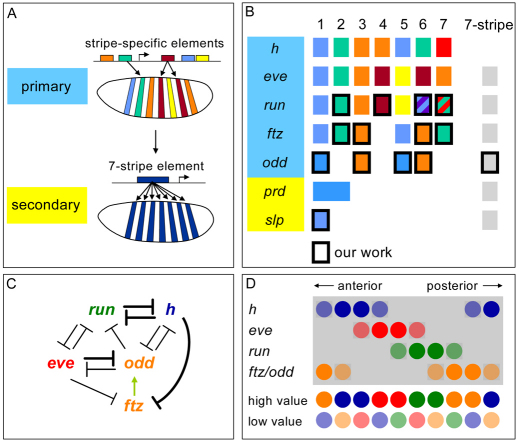
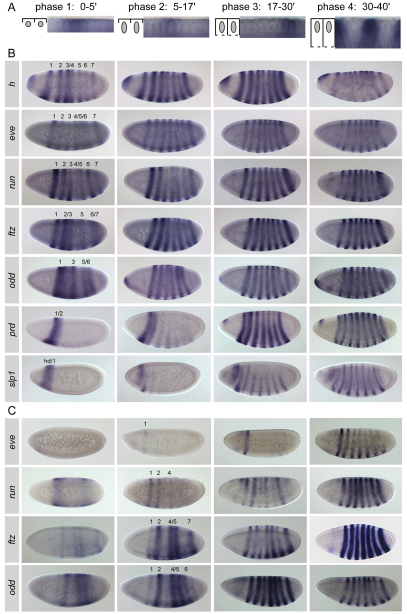
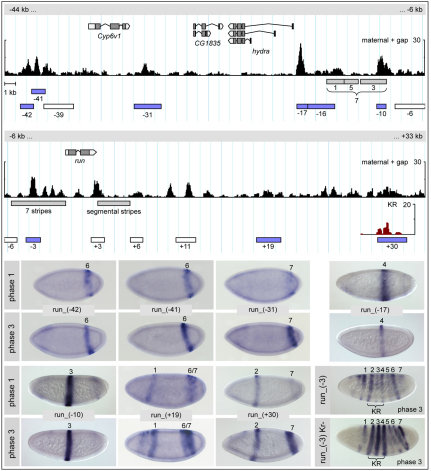
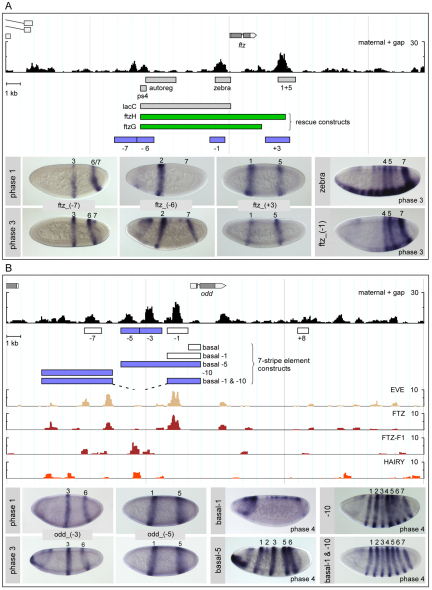
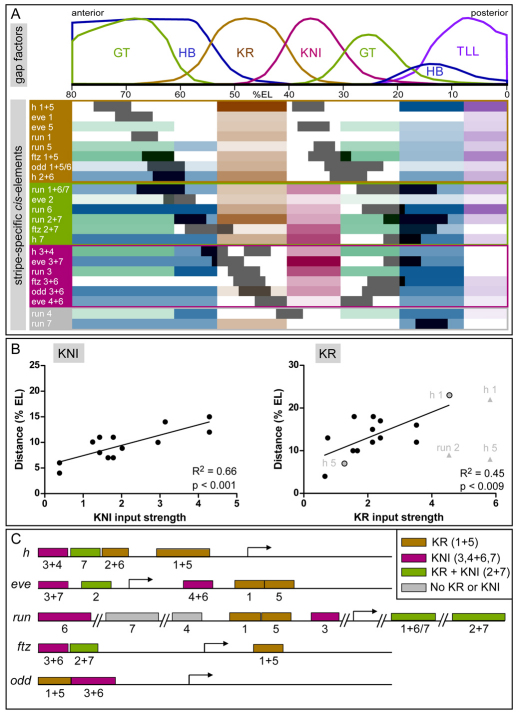
The generation of metameric body plans is a key process in development. In Drosophila segmentation, periodicity is established rapidly through the complex transcriptional regulation of the pair-rule genes. The 'primary' pair-rule genes generate their 7-stripe expression through stripe-specific cis-regulatory elements controlled by the preceding non-periodic maternal and gap gene patterns, whereas 'secondary' pair-rule genes are thought to rely on 7-stripe elements that read off the already periodic primary pair-rule patterns. Using a combination of computational and experimental approaches, we have conducted a comprehensive systems-level examination of the regulatory architecture underlying pair-rule stripe formation. We find that runt (run), fushi tarazu (ftz) and odd skipped (odd) establish most of their pattern through stripe-specific elements, arguing for a reclassification of ftz and odd as primary pair-rule genes. In the case of run, we observe long-range cis-regulation across multiple intervening genes. The 7-stripe elements of run, ftz and odd are active concurrently with the stripe-specific elements, indicating that maternal/gap-mediated control and pair-rule gene cross-regulation are closely integrated. Stripe-specific elements fall into three distinct classes based on their principal repressive gap factor input; stripe positions along the gap gradients correlate with the strength of predicted input. The prevalence of cis-elements that generate two stripes and their genomic organization suggest that single-stripe elements arose by splitting and subfunctionalization of ancestral dual-stripe elements. Overall, our study provides a greatly improved understanding of how periodic patterns are established in the Drosophila embryo.
Figures

Similar articles
-
Huckebein is part of a combinatorial repression code in the anterior blastoderm.Dev Biol. 2012 Jan 1;361(1):177-85. doi: 10.1016/j.ydbio.2011.10.016. Epub 2011 Oct 15. Dev Biol. 2012. PMID: 22027434
-
Modelling and calibration of pair-rule protein patterns in Drosophila embryo: From Even-skipped and Fushi-tarazu to Wingless expression networks.Dev Biol. 2025 Jan;517:178-190. doi: 10.1016/j.ydbio.2024.09.015. Epub 2024 Oct 4. Dev Biol. 2025. PMID: 39369936
-
Different modes of enhancer-specific regulation by Runt and Even-skipped during Drosophila segmentation.Mol Biol Cell. 2017 Mar 1;28(5):681-691. doi: 10.1091/mbc.E16-09-0630. Epub 2017 Jan 11. Mol Biol Cell. 2017. PMID: 28077616 Free PMC article.
-
Transcriptional control by Drosophila gap genes.J Cell Sci Suppl. 1992;16:39-51. doi: 10.1242/jcs.1992.supplement_16.6. J Cell Sci Suppl. 1992. PMID: 1297651 Review.
-
Modeling the Drosophila pair-rule pattern by reaction-diffusion: gap input and pattern control in a 4-morphogen system.J Theor Biol. 1990 May 22;144(2):171-94. doi: 10.1016/s0022-5193(05)80317-0. J Theor Biol. 1990. PMID: 2197508 Review.
Cited by
-
Transcriptional Enhancers in Drosophila.Genetics. 2020 Sep;216(1):1-26. doi: 10.1534/genetics.120.301370. Genetics. 2020. PMID: 32878914 Free PMC article. Review.
-
A Mutation in the Drosophila melanogaster eve Stripe 2 Minimal Enhancer Is Buffered by Flanking Sequences.G3 (Bethesda). 2020 Dec 3;10(12):4473-4482. doi: 10.1534/g3.120.401777. G3 (Bethesda). 2020. PMID: 33037064 Free PMC article.
-
Expression of pair rule gene orthologs in the blastoderm of a myriapod: evidence for pair rule-like mechanisms?BMC Dev Biol. 2012 May 17;12:15. doi: 10.1186/1471-213X-12-15. BMC Dev Biol. 2012. PMID: 22595029 Free PMC article.
-
Mechanisms of Interaction between Enhancers and Promoters in Three Drosophila Model Systems.Int J Mol Sci. 2023 Feb 2;24(3):2855. doi: 10.3390/ijms24032855. Int J Mol Sci. 2023. PMID: 36769179 Free PMC article. Review.
-
Dynamic modulation of enhancer responsiveness by core promoter elements in living Drosophila embryos.Nucleic Acids Res. 2022 Jan 11;50(1):92-107. doi: 10.1093/nar/gkab1177. Nucleic Acids Res. 2022. PMID: 34897508 Free PMC article.
References
-
- Akam M. (1987). The molecular basis for metameric pattern in the Drosophila embryo. Development 101, 1-22 - PubMed
-
- Andrioli L. P., Oberstein A. L., Corado M. S., Yu D., Small S. (2004). Groucho-dependent repression by sloppy-paired 1 differentially positions anterior pair-rule stripes in the Drosophila embryo. Dev. Biol. 276, 541-551 - PubMed
-
- Arnosti D. N., Barolo S., Levine M., Small S. (1996a). The eve stripe 2 enhancer employs multiple modes of transcriptional synergy. Development 122, 205-214 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
