

Review
doi: 10.1021/cr200075y.
Epub 2011 Jun 23.
The metabolic serine hydrolases and their functions in mammalian physiology and disease
Affiliations
- PMID: 21696217
- PMCID: PMC3192302
- DOI: 10.1021/cr200075y
Item in Clipboard
Review
The metabolic serine hydrolases and their functions in mammalian physiology and disease
Chem Rev.
.
No abstract available
Figures
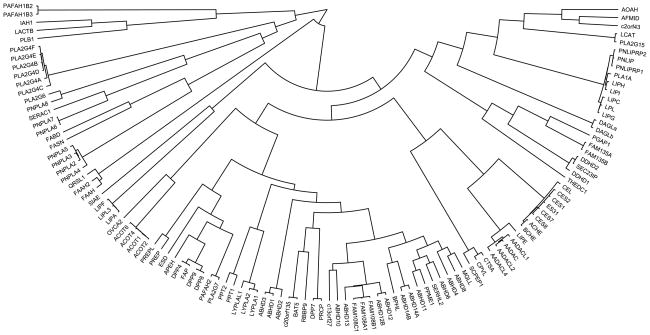
Dendrogram showing the primary sequence alignment of the human mammalian metabolic SHs, where alignment was generated by anchoring sequences at the site of their catalytic Ser residues.
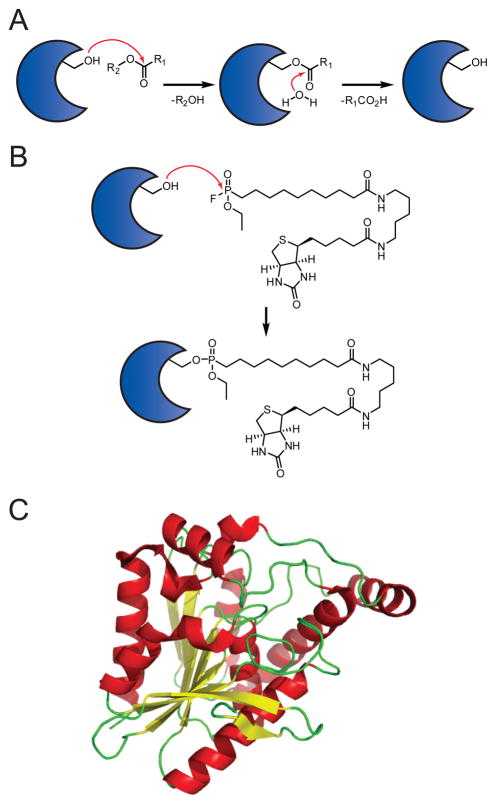
(A) Mechanism of SH catalysis. (B) Mechanism of SH labeling by the active site-directed activity-based probe fluorophosphonate-biotin (FP-biotin). (C) Three dimensional structure of MGLL, a SH with a canonical α/β-hydrolase fold.

The enzymatic catabolism of triglycerides into fatty acids and glycerol by PNPLA2, HSL, DAGLα/β, and MGLL.
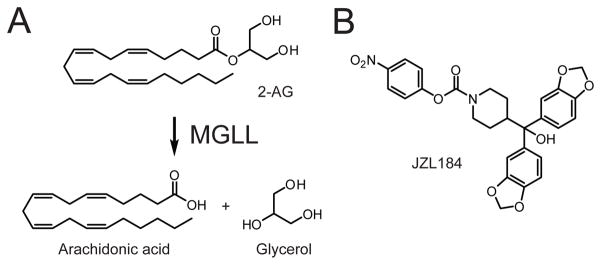
(A) MGLL hydrolyzes the endocannabinoid 2-arachidonoylglycerol (2-AG) to generate arachidonic acid and glycerol. (B) Structure of the MGLL-selective inhibitor JZL184.
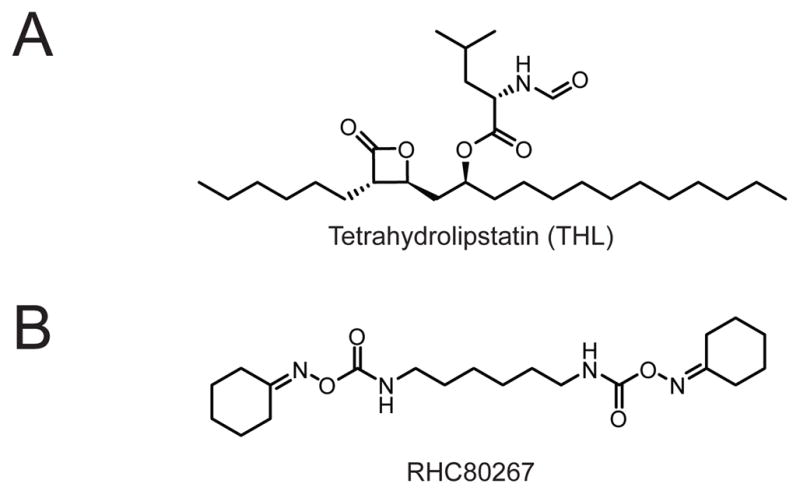
(A) Structure of the non-selective lipase inhibitor tetrahydrolipstatin (THL, also called Orlistat, Xenical, or Alli). (B) Structure of the non-selective DAGL inhibitor RHC80267.
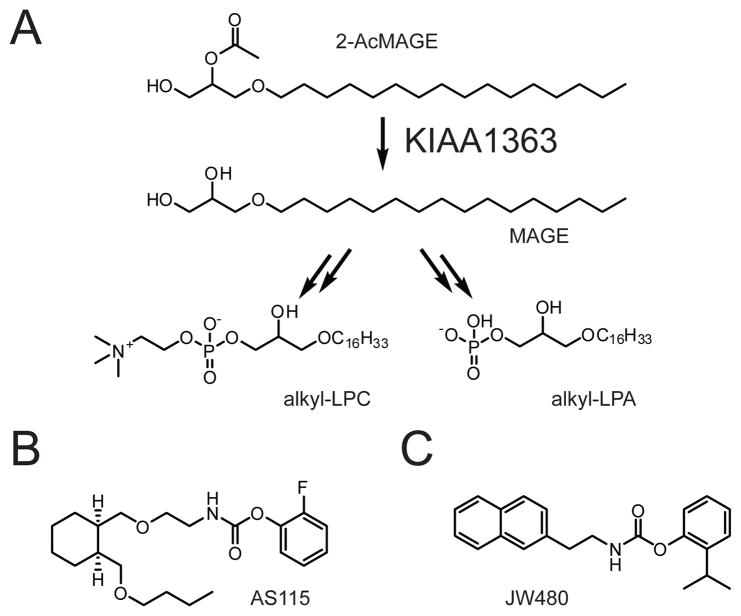
(A) KIAA1363 hydrolyzes 2-AcMAGE to generate MAGE, which can be further converted to alkyl-LPC (left) or alkyl-LPA (right). (B, C) Structures of the lead KIAA1363 inhibitor AS115 (B) and the more advanced KIAA1363 inhibitor JW480 (C), which shows improved selectivity and in vivo activity.

(A, B) Structures of the ABHD6 inhibitors WWL70 (A) and WWL123 (B).
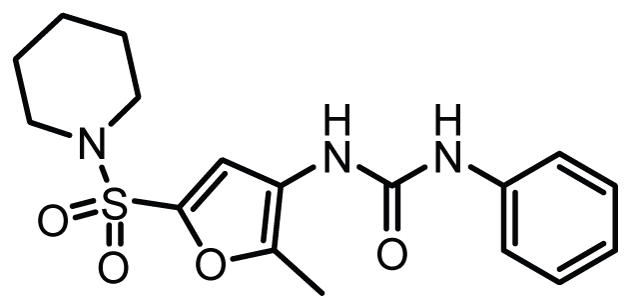
Structure of a urea-based dual LPL/LIPG inhibitor.
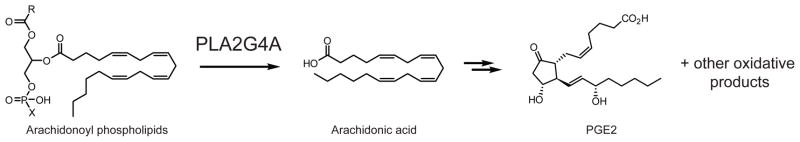
PLA2G4A liberates arachidonic acid from sn-2 arachidonoyl-containing phospholipids in macrophages. The arachidonate generated by this reaction can be further oxidized by cyclooxygenases, lipoxygenases, and prostaglandin synthases into eicosanoid such as PGE2 (shown) and other oxidative derivatives.

Structures of indole-derived PLA2G4A inhibitors.
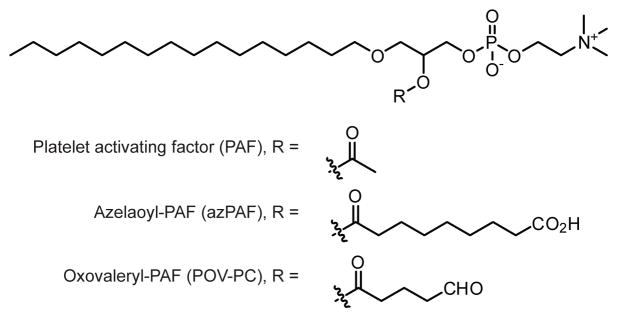
Structures of oxidatively-truncated phosphatidylcholine species.
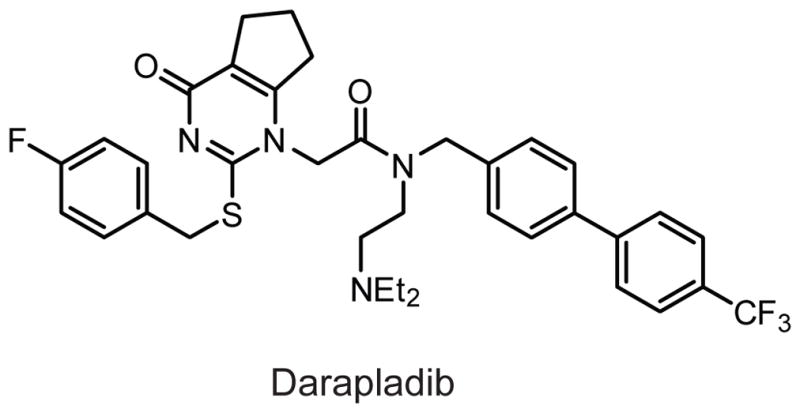
Structure of the PLA2G7 inhibitor darapladib.
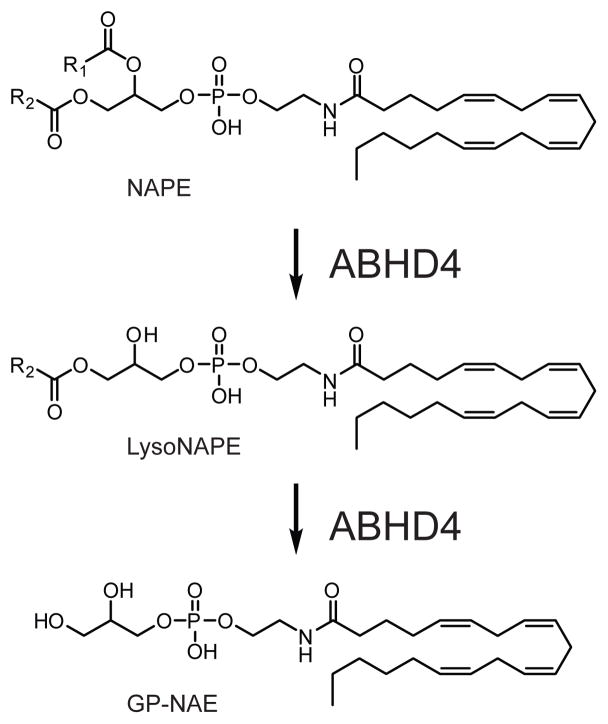
ABHD4 sequentially hydrolyzes O-acyl chains from NAPEs to generate lysoNAPEs and glycerophospho-NAEs (GP-NAEs).
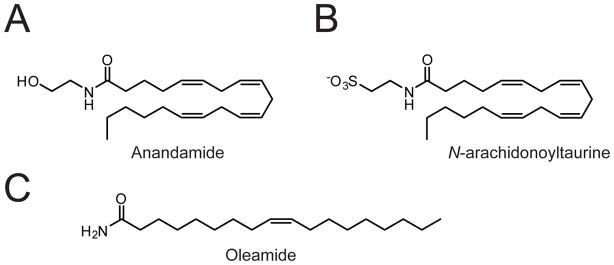
(A-C) Structure of the FAAH substrates anandamide (A), N-arachidonoyltaurine (B), and oleamide (C).
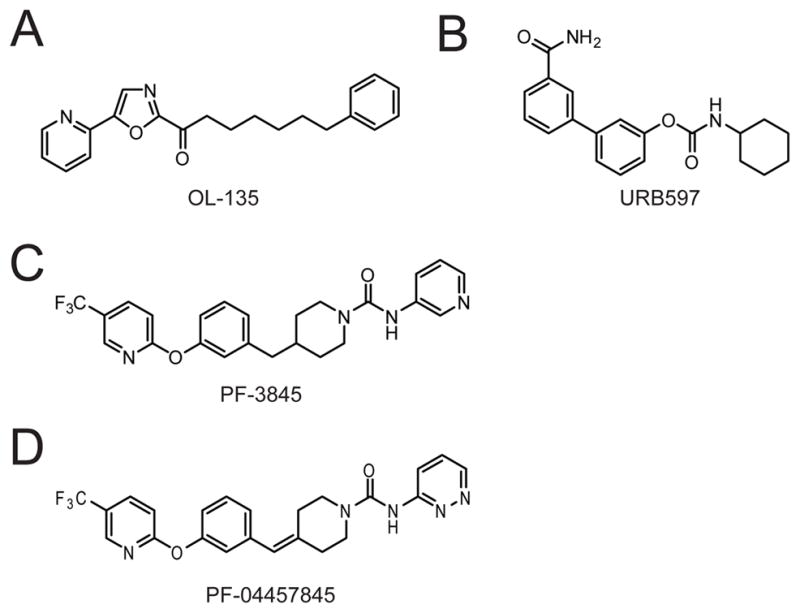
(A-D) Structure of the FAAH inhibitors OL-135 (A), URB597 (B), PF-3845 (C), and PF-04457845 (D).
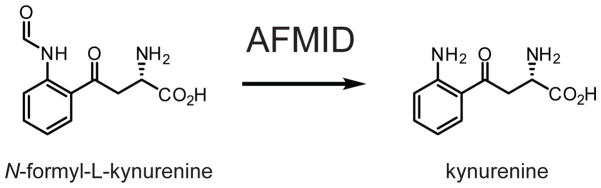
AFMID hydrolyzes N-formyl-L-kynurenine to generate kynurenine.
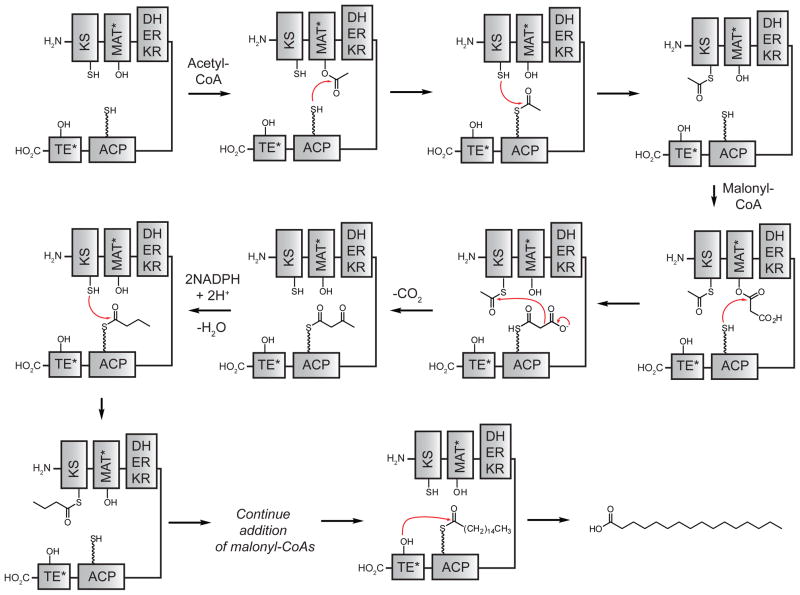
The FASN catalytic cycle. The seven domains are abbreviated β-ketoacyl synthase (KS), malonyl-CoA-/acetyl-CoA-ACP-transacylase (MAT), dehydratase (DH), β-enoyl reductase (ER), β-ketoacyl reductase (KR), acyl carrier protein (ACP), and thioesterase (TE). The residues are abbreviated –OH (serine), –SH (cysteine), and
 SH (4′-phosphopantetheine). MAT* and TE* indicate that these two are SH domains. Following attachment of the initial acetate, the cycle is repeated seven times to generate one equivalent of palmitic acid.
SH (4′-phosphopantetheine). MAT* and TE* indicate that these two are SH domains. Following attachment of the initial acetate, the cycle is repeated seven times to generate one equivalent of palmitic acid.
SH (4′-phosphopantetheine). MAT* and TE* indicate that these two are SH domains. Following attachment of the initial acetate, the cycle is repeated seven times to generate one equivalent of palmitic acid.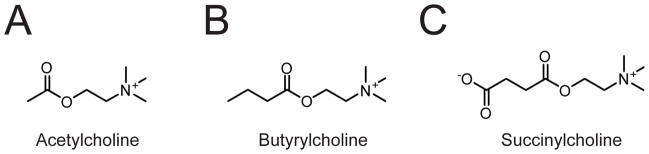
(A-C) Structure of the ACHE substrate acetylcholine (A) and of the BCHE substrates butyrylcholine (B) and succinylcholine (C).
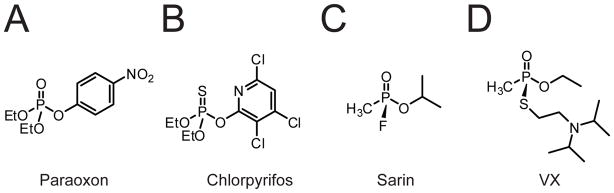
(A-D) Structures of the organophosphorous insecticides paraoxon (A) and chlorpyrifos (B), and the nerve agents sarin (C) and VX (D).
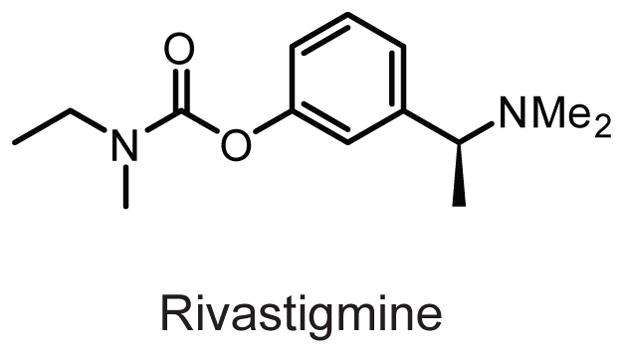
Structure of the dual ACHE/BCHE inhibitor rivastigmine.
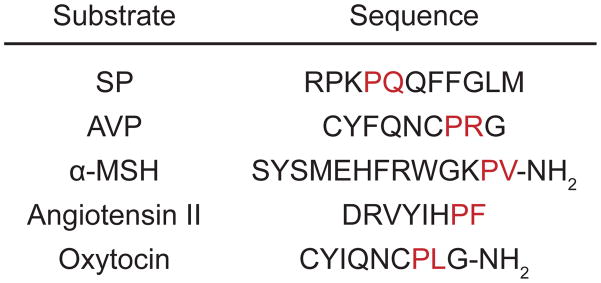
Cleavage positions of some representative PREP substrates, with the scissile bond between the amino acids shown in red. SP, substance P; AVP, arginine-vasopressin; α-MSH, α-melanocyte stimulating hormone.
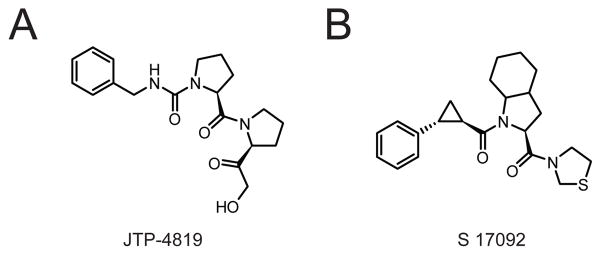
(A, B) Structures of the PREP inhibitors JTP-4819 (A) and S 17092 (B).

Structure of GLP-1, a representative substrate of DPP4, which is cleaved to liberate an N-terminal dipeptide (cleavage site shown in red). Other DPP4 substrates are also cleaved at the same penultimate position.
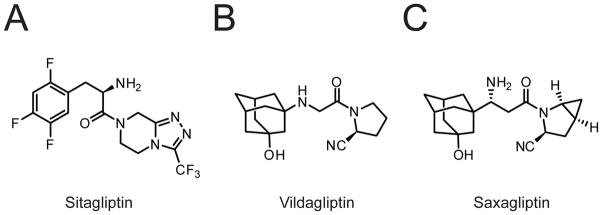
The DPP4 inhibitors sitagliptin (A, also called Januvia), vildagliptin (B), and saxagliptin (C).
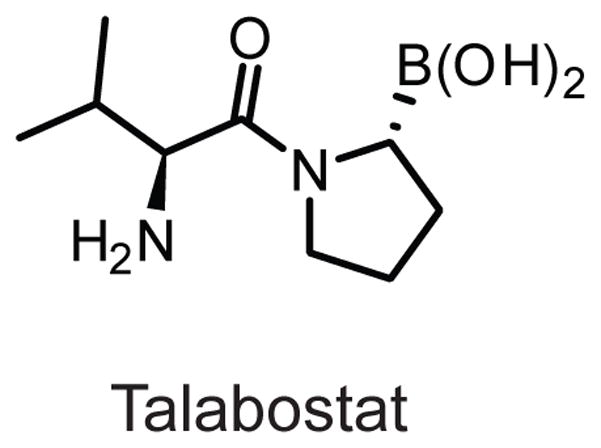
Structure of the non-selective DPP-family inhibitor talabostat.

APEH cleaves N-acetylated peptides to generate an N-acetyl amino acid and an unblocked peptide.
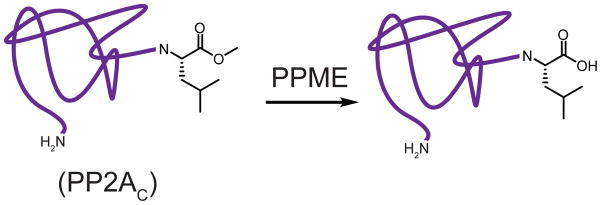
PPME hydrolyzes the carboxymethylated C-terminal leucine residue of the PP2A catalytic subunit (PP2AC). Shown in purple, PP2AC.
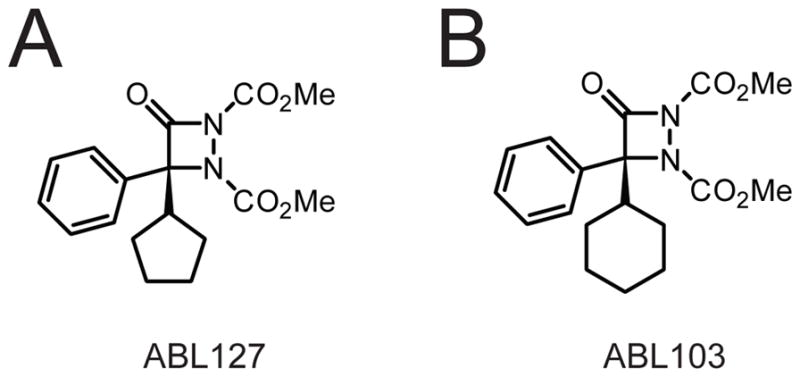
(A, B) Structure of the aza-β-lactam-based PPME inhibitors ABL127 (A) and ABL103 (B).
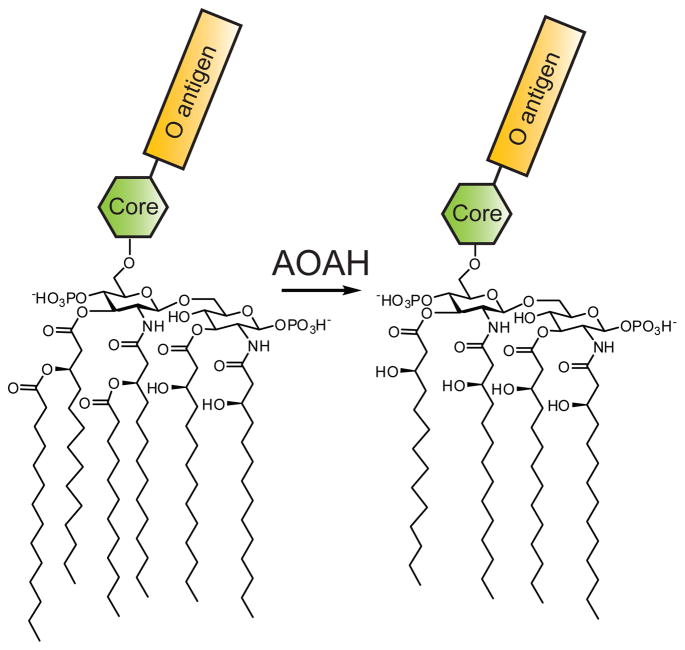
AOAH hydrolyzes the two acyloxyacyl groups from the lipid A portion of LPS to generate tetraacyl-LPS. The core oligosaccharide and O antigen of LPS are represented by the green hexagon and yellow rectangle, respectively.
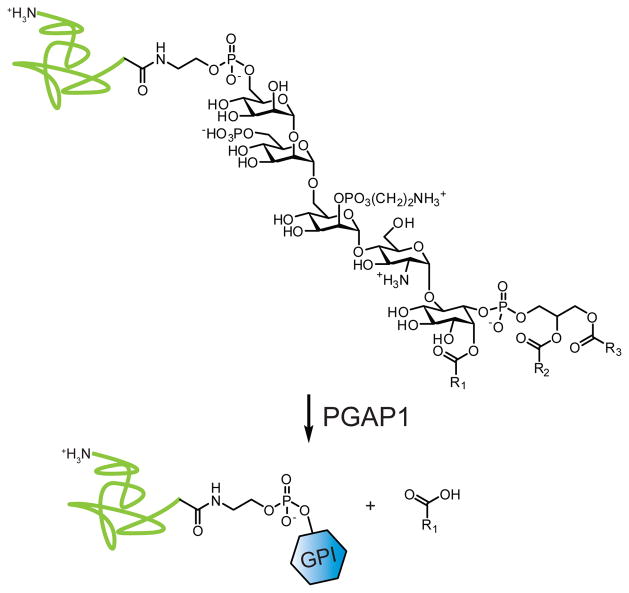
PGAP1 hydrolyzes palmitate from the inositol of triacyl GPI to generate mature, diacyl-GPI. Shown in green, a protein modified by GPI; shown as a blue hexagon, a mature diacyl-GPI modification.
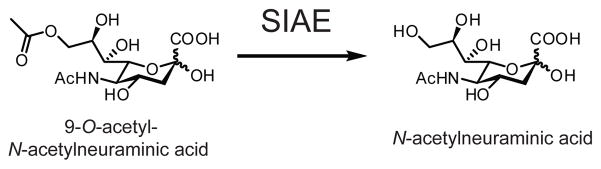
SIAE removes acetate from 9-O-acetyl-N-acetylneuraminic acid (shown), as well as other 9-O-acetylated sialic acids. This activity can occur on both free acetylated sialic acids or on acetylated sialic acid found in glycoproteins.
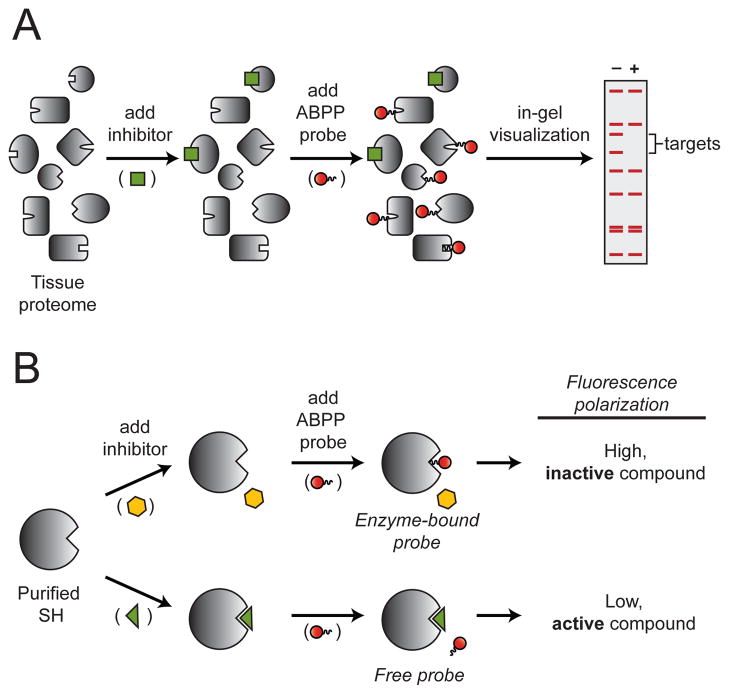
(A) Competitive ABPP. The potency and selectivity of a single inhibitor (green square) can be evaluated against SHs in a native tissue proteome by chasing the inhibitor with a SH-directed activity probe (red circle) and visualization by in-gel fluorescence following SDS-PAGE separation. Inhibitor targets are detected as bands with reduced fluorescence. (B) Fluopol-ABPP. A single SH can be evaluated against many compounds by chasing inhibitors (yellow hexagon or green triangle) with a SH-directed activity probe (red circle). Active (green triangle) or inactive (yellow hexagon) inhibitors will produce low or high fluorescence polarization signals, respectively.
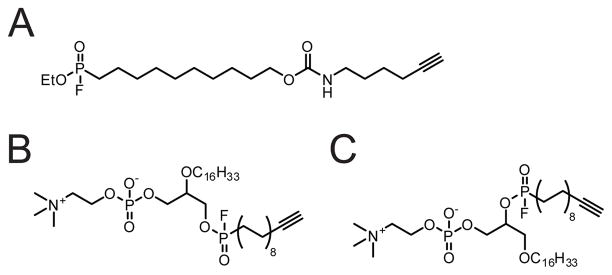
(A) Structure of the general SH-directed probe FP-alkyne. (B, C) Structure of the sn-1-selective (B) or sn-2-selective (C) phospholipase-directed probes.
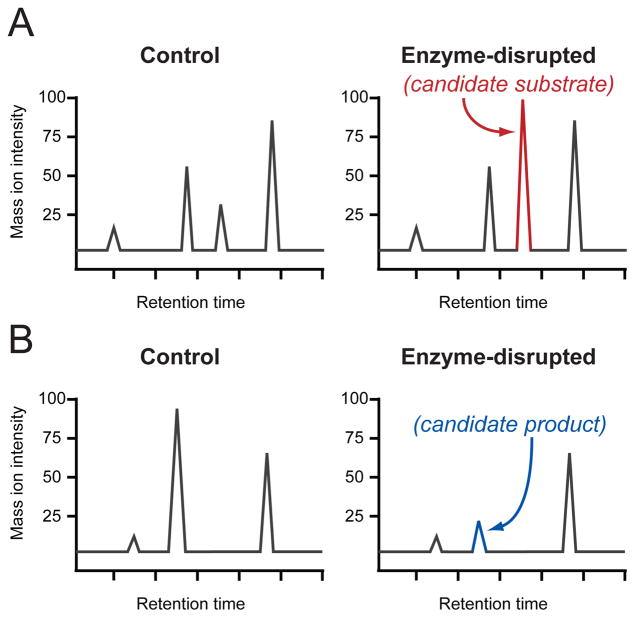
DMP can identify metabolite peaks in LC-MS analyses with relative levels that are either increasing (A) or decreasing (B) between control and enzyme-disrupted samples. The increasing or decreasing peaks correspond to candidate substrates and products, respectively. Standard-free, relative quantitation is achieved by comparison to the many other peaks that are not changing (shown in grey).
References
-
- Powers JC, Asgian JL, Ekici OD, James KE. Chem Rev. 2002;102:4639. - PubMed
-
- Barrett AJ, Rawlings ND, Woessner JF, editors. Handbook of Proteolytic Enzymes. Academic Press; Bath: 1998.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
