

Impact of epistasis and pleiotropy on evolutionary adaptation
- PMID: 21697174
- PMCID: PMC3223680
- DOI: 10.1098/rspb.2011.0870
Impact of epistasis and pleiotropy on evolutionary adaptation
Abstract
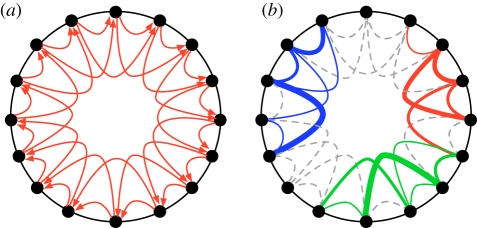
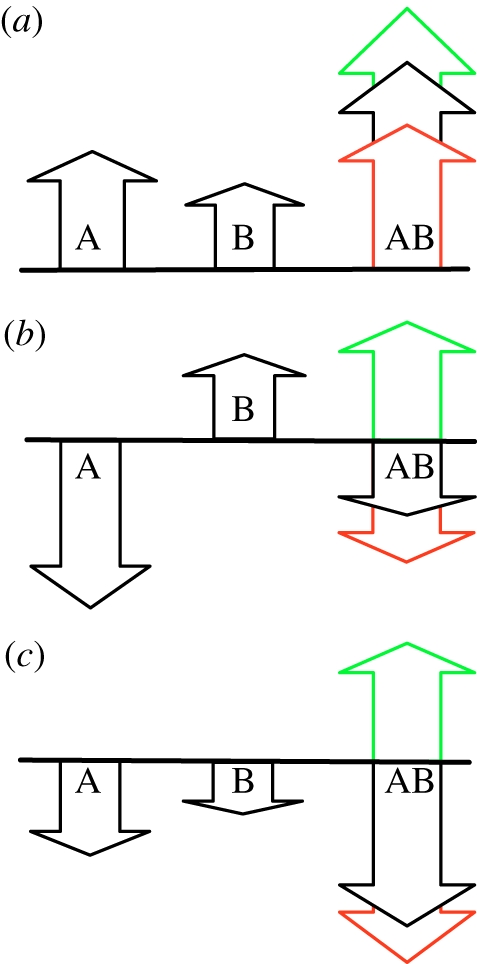
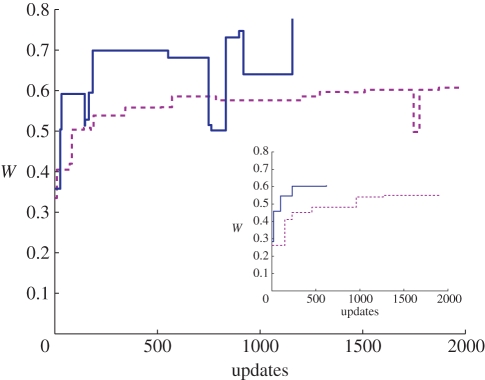
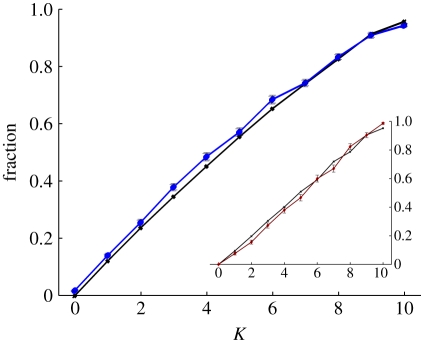
Evolutionary adaptation is often likened to climbing a hill or peak. While this process is simple for fitness landscapes where mutations are independent, the interaction between mutations (epistasis) as well as mutations at loci that affect more than one trait (pleiotropy) are crucial in complex and realistic fitness landscapes. We investigate the impact of epistasis and pleiotropy on adaptive evolution by studying the evolution of a population of asexual haploid organisms (haplotypes) in a model of N interacting loci, where each locus interacts with K other loci. We use a quantitative measure of the magnitude of epistatic interactions between substitutions, and find that it is an increasing function of K. When haplotypes adapt at high mutation rates, more epistatic pairs of substitutions are observed on the line of descent than expected. The highest fitness is attained in landscapes with an intermediate amount of ruggedness that balance the higher fitness potential of interacting genes with their concomitant decreased evolvability. Our findings imply that the synergism between loci that interact epistatically is crucial for evolving genetic modules with high fitness, while too much ruggedness stalls the adaptive process.
Figures
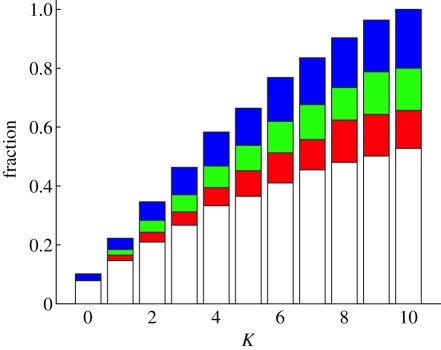
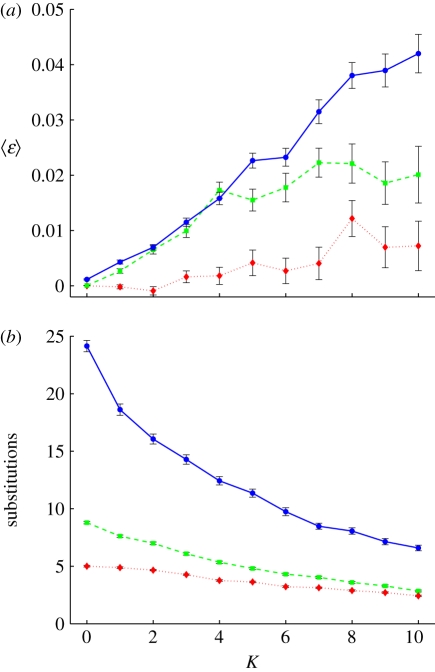
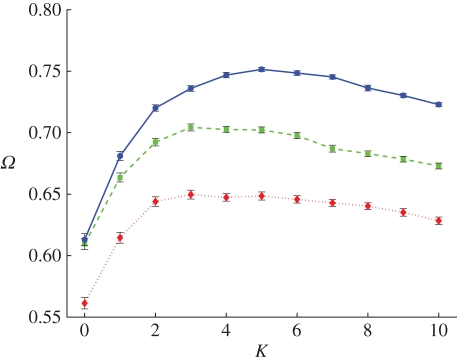
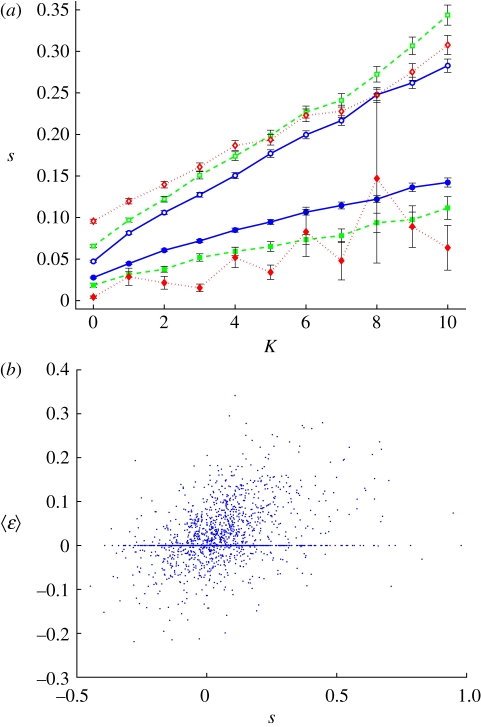
References
-
- Fisher R. 1930. The genetical theory of natural selection. Oxford, UK: Oxford University Press
-
- Orr H. A. 2005. The genetic theory of adaptation: a brief history. Nat. Rev. Genet. 6, 119–12710.1038/nrg1523 (doi:10.1038/nrg1523) - DOI - DOI - PubMed
-
- Gillespie J. H. 1984. Molecular evolution over the mutational landscape. Evolution 38, 1116–112910.2307/2408444 (doi:10.2307/2408444) - DOI - DOI - PubMed
-
- Gillespie J. 1991. The causes of molecular evolution. New York, NY: Oxford University Press
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources