

Functional correlates of the anterolateral processing hierarchy in human auditory cortex
- PMID: 21697384
- PMCID: PMC3142575
- DOI: 10.1523/JNEUROSCI.1448-11.2011
Functional correlates of the anterolateral processing hierarchy in human auditory cortex
Abstract
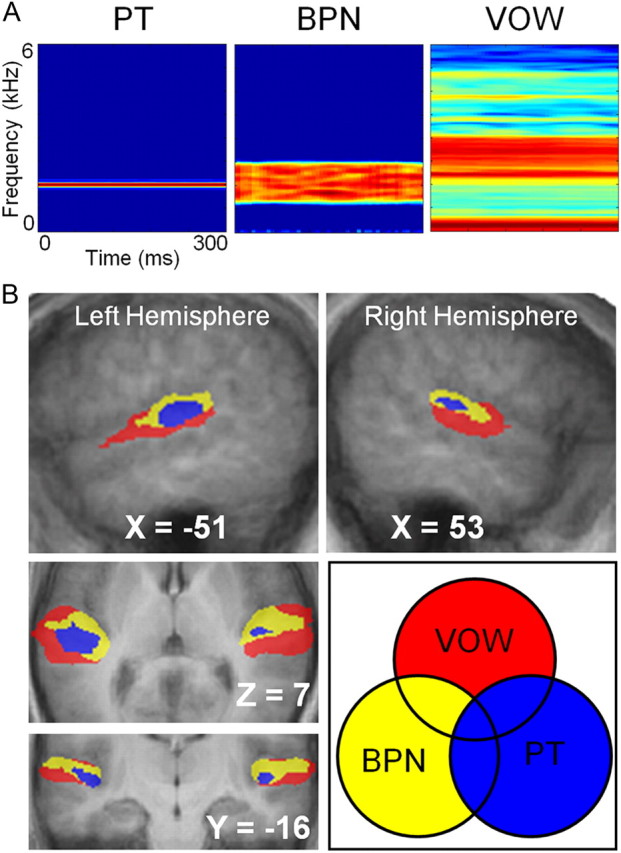
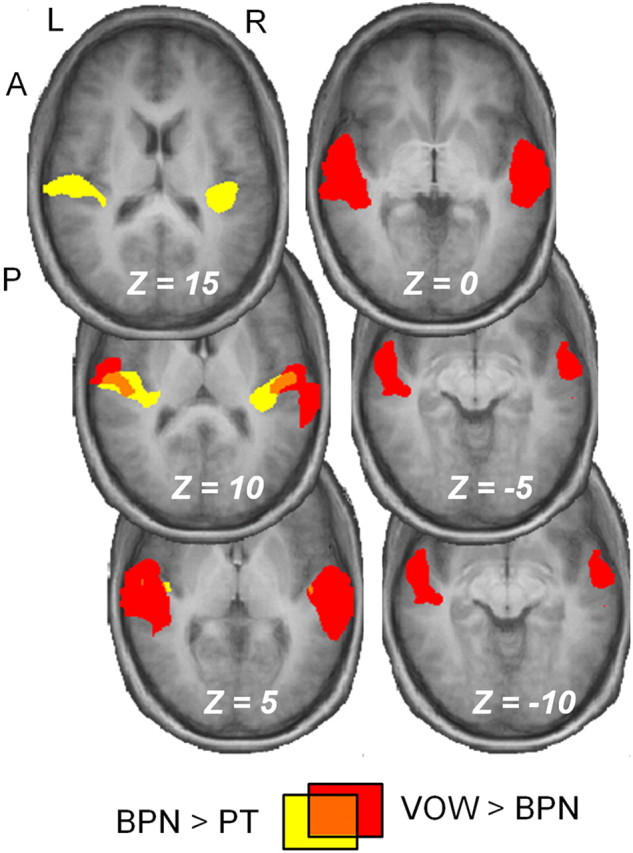
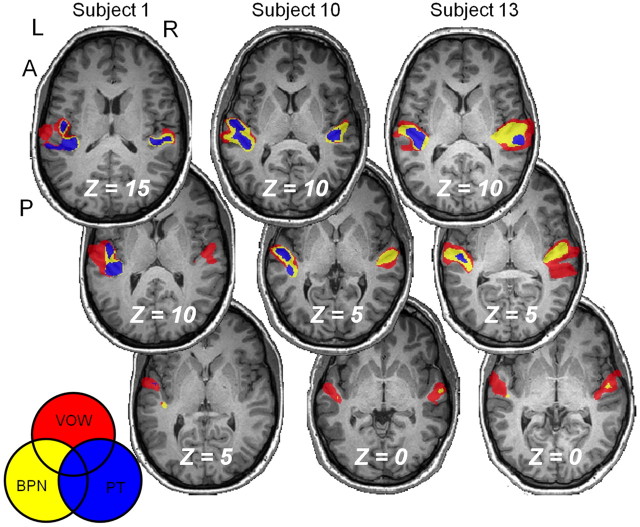
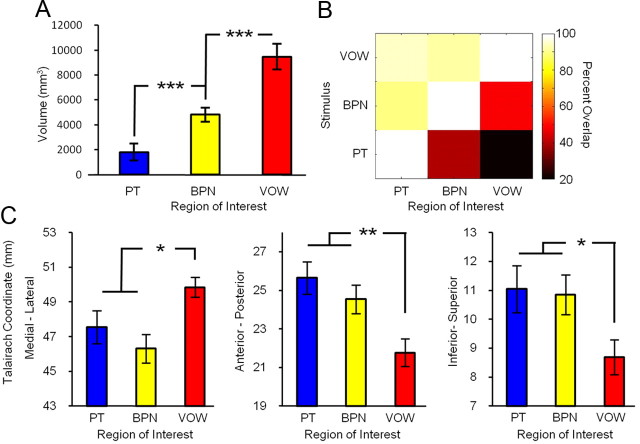
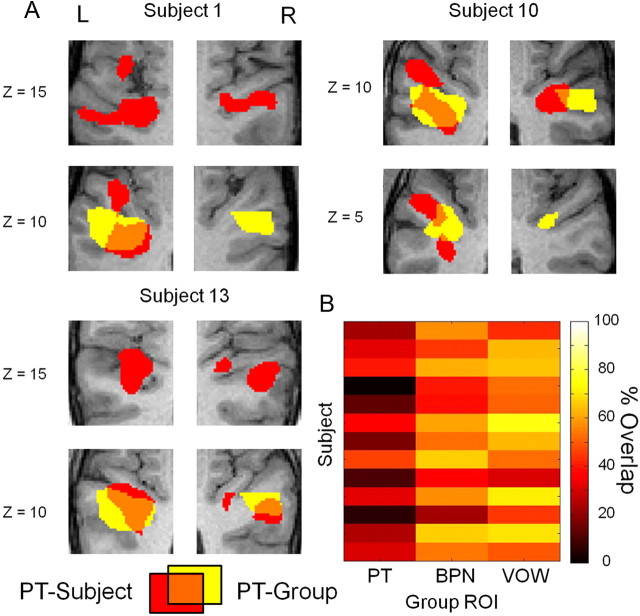
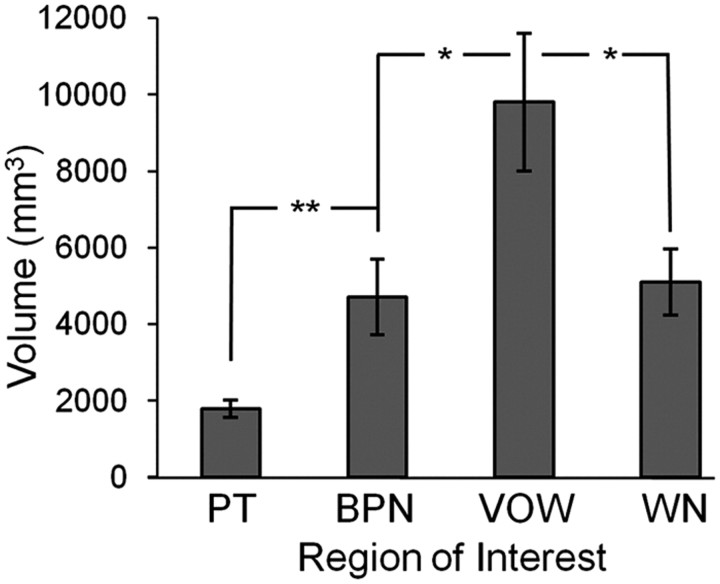
Converging evidence supports the hypothesis that an anterolateral processing pathway mediates sound identification in auditory cortex, analogous to the role of the ventral cortical pathway in visual object recognition. Studies in nonhuman primates have characterized the anterolateral auditory pathway as a processing hierarchy, composed of three anatomically and physiologically distinct initial stages: core, belt, and parabelt. In humans, potential homologs of these regions have been identified anatomically, but reliable and complete functional distinctions between them have yet to be established. Because the anatomical locations of these fields vary across subjects, investigations of potential homologs between monkeys and humans require these fields to be defined in single subjects. Using functional MRI, we presented three classes of sounds (tones, band-passed noise bursts, and conspecific vocalizations), equivalent to those used in previous monkey studies. In each individual subject, three regions showing functional similarities to macaque core, belt, and parabelt were readily identified. Furthermore, the relative sizes and locations of these regions were consistent with those reported in human anatomical studies. Our results demonstrate that the functional organization of the anterolateral processing pathway in humans is largely consistent with that of nonhuman primates. Because our scanning sessions last only 15 min/subject, they can be run in conjunction with other scans. This will enable future studies to characterize functional modules in human auditory cortex at a level of detail previously possible only in visual cortex. Furthermore, the approach of using identical schemes in both humans and monkeys will aid with establishing potential homologies between them.
Figures
Similar articles
-
Functional organization of human auditory cortex: investigation of response latencies through direct recordings.Neuroimage. 2014 Nov 1;101:598-609. doi: 10.1016/j.neuroimage.2014.07.004. Epub 2014 Jul 12. Neuroimage. 2014. PMID: 25019680 Free PMC article.
-
Sound Frequency Representation in the Auditory Cortex of the Common Marmoset Visualized Using Optical Intrinsic Signal Imaging.eNeuro. 2018 May 7;5(2):ENEURO.0078-18.2018. doi: 10.1523/ENEURO.0078-18.2018. eCollection 2018 Mar-Apr. eNeuro. 2018. PMID: 29736410 Free PMC article.
-
A hierarchy of processing complexity and timescales for natural sounds in the human auditory cortex.Proc Natl Acad Sci U S A. 2025 May 6;122(18):e2412243122. doi: 10.1073/pnas.2412243122. Epub 2025 Apr 28. Proc Natl Acad Sci U S A. 2025. PMID: 40294254
-
Processing of complex sounds in the auditory cortex of cat, monkey, and man.Acta Otolaryngol Suppl. 1997;532:34-8. doi: 10.3109/00016489709126142. Acta Otolaryngol Suppl. 1997. PMID: 9442842 Review.
-
Subdivisions of auditory cortex and levels of processing in primates.Audiol Neurootol. 1998 Mar-Jun;3(2-3):73-85. doi: 10.1159/000013783. Audiol Neurootol. 1998. PMID: 9575378 Review.
Cited by
-
A high-resolution 7-Tesla fMRI dataset from complex natural stimulation with an audio movie.Sci Data. 2014 May 27;1:140003. doi: 10.1038/sdata.2014.3. eCollection 2014. Sci Data. 2014. PMID: 25977761 Free PMC article.
-
Exploring Hierarchical Auditory Representation via a Neural Encoding Model.Front Neurosci. 2022 Mar 24;16:843988. doi: 10.3389/fnins.2022.843988. eCollection 2022. Front Neurosci. 2022. PMID: 35401085 Free PMC article.
-
Abnormal auditory tonotopy in patients with schizophrenia.NPJ Schizophr. 2019 Oct 2;5(1):16. doi: 10.1038/s41537-019-0084-x. NPJ Schizophr. 2019. PMID: 31578332 Free PMC article.
-
Selectivity for space and time in early areas of the auditory dorsal stream in the rhesus monkey.J Neurophysiol. 2014 Apr;111(8):1671-85. doi: 10.1152/jn.00436.2013. Epub 2014 Feb 5. J Neurophysiol. 2014. PMID: 24501260 Free PMC article.
-
Parcellation of Human and Monkey Core Auditory Cortex with fMRI Pattern Classification and Objective Detection of Tonotopic Gradient Reversals.Cereb Cortex. 2015 Oct;25(10):3278-89. doi: 10.1093/cercor/bhu124. Epub 2014 Jun 5. Cereb Cortex. 2015. PMID: 24904067 Free PMC article.
References
-
- Belin P, Zatorre RJ, Lafaille P, Ahad P, Pike B. Voice-selective areas in human auditory cortex. Nature. 2000;403:309–312. - PubMed
-
- Binder JR, Frost JA, Hammeke TA, Bellgowan PS, Springer JA, Kaufman JN, Possing ET. Human temporal lobe activation by speech and nonspeech sounds. Cereb Cortex. 2000;10:512–528. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources