

Genome-wide function of THO/TREX in active genes prevents R-loop-dependent replication obstacles
- PMID: 21701562
- PMCID: PMC3160181
- DOI: 10.1038/emboj.2011.206
Genome-wide function of THO/TREX in active genes prevents R-loop-dependent replication obstacles
Abstract
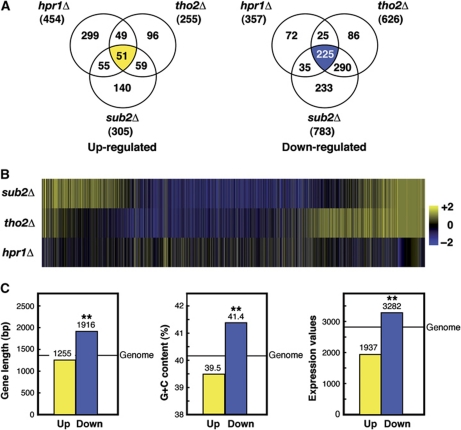
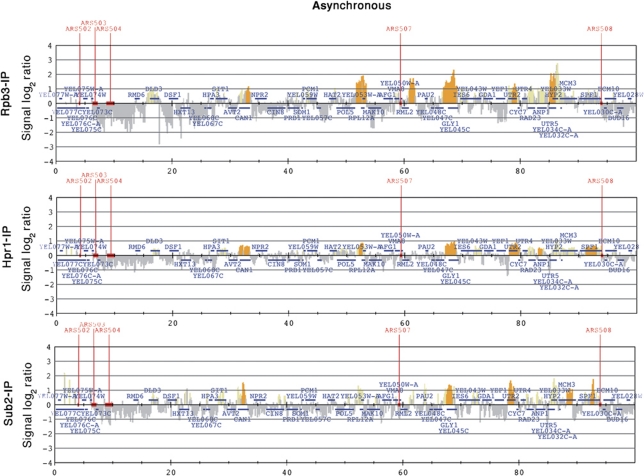
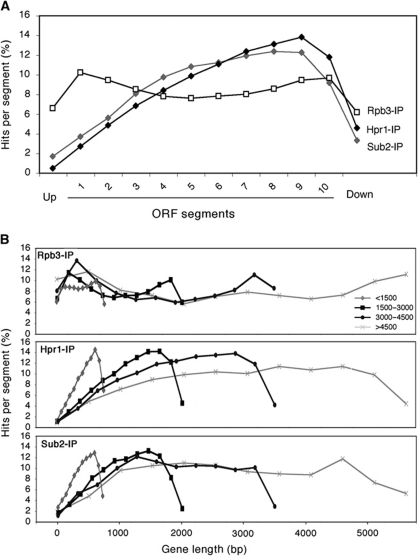
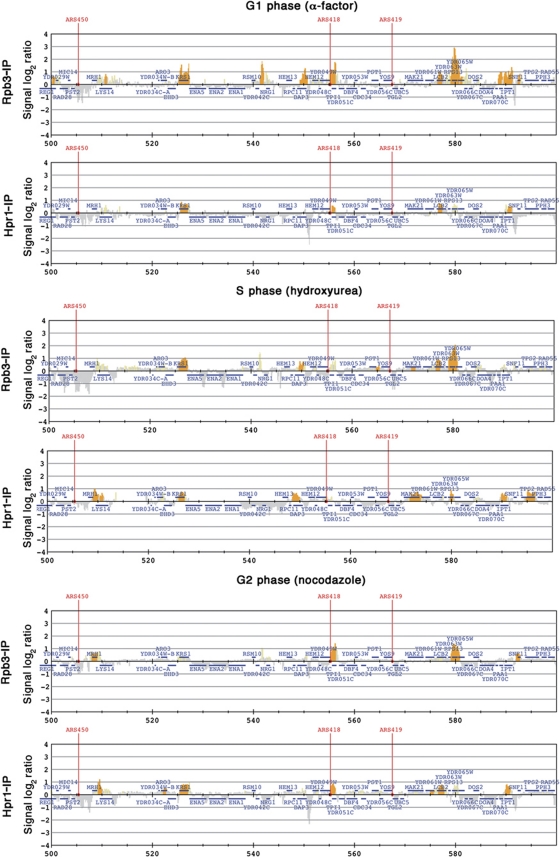
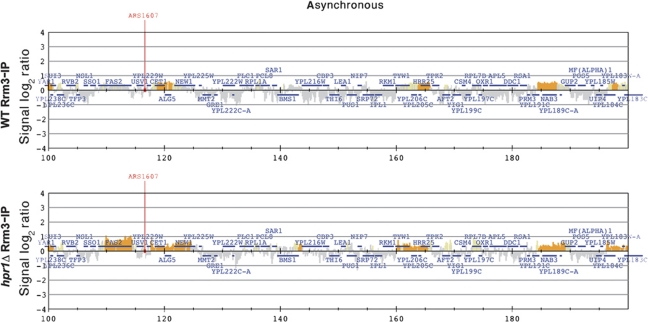
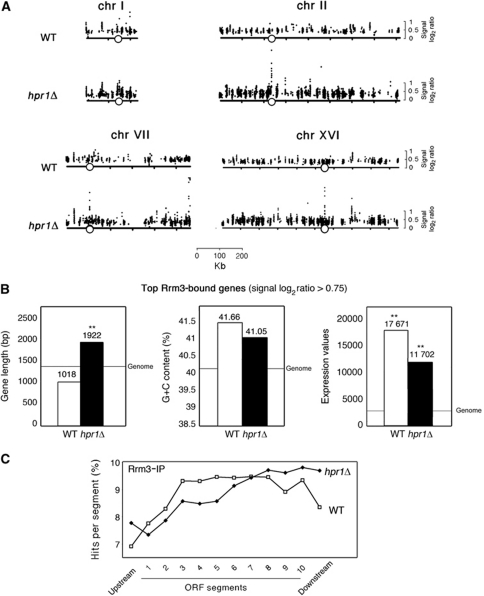
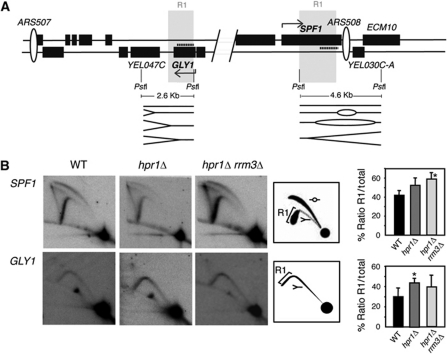
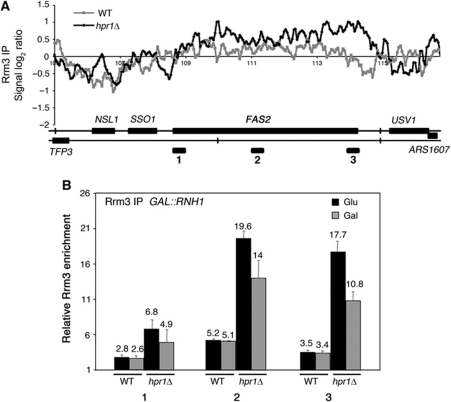
THO/TREX is a conserved nuclear complex that functions in mRNP biogenesis and prevents transcription-associated recombination. Whether or not it has a ubiquitous role in the genome is unknown. Chromatin immunoprecipitation (ChIP)-chip studies reveal that the Hpr1 component of THO and the Sub2 RNA-dependent ATPase have genome-wide distributions at active ORFs in yeast. In contrast to RNA polymerase II, evenly distributed from promoter to termination regions, THO and Sub2 are absent at promoters and distributed in a gradual 5' → 3' gradient. This is accompanied by a genome-wide impact of THO-Sub2 deletions on expression of highly expressed, long and high G+C-content genes. Importantly, ChIP-chips reveal an over-recruitment of Rrm3 in active genes in THO mutants that is reduced by RNaseH1 overexpression. Our work establishes a genome-wide function for THO-Sub2 in transcription elongation and mRNP biogenesis that function to prevent the accumulation of transcription-mediated replication obstacles, including R-loops.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
