

Divergent activities of osteogenic BMP2, and tenogenic BMP12 and BMP13 independent of receptor binding affinities
- PMID: 21702718
- PMCID: PMC3154542
- DOI: 10.3109/08977194.2011.593178
Divergent activities of osteogenic BMP2, and tenogenic BMP12 and BMP13 independent of receptor binding affinities
Abstract
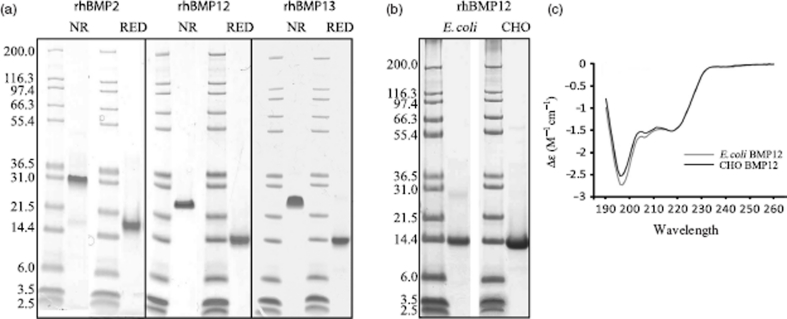
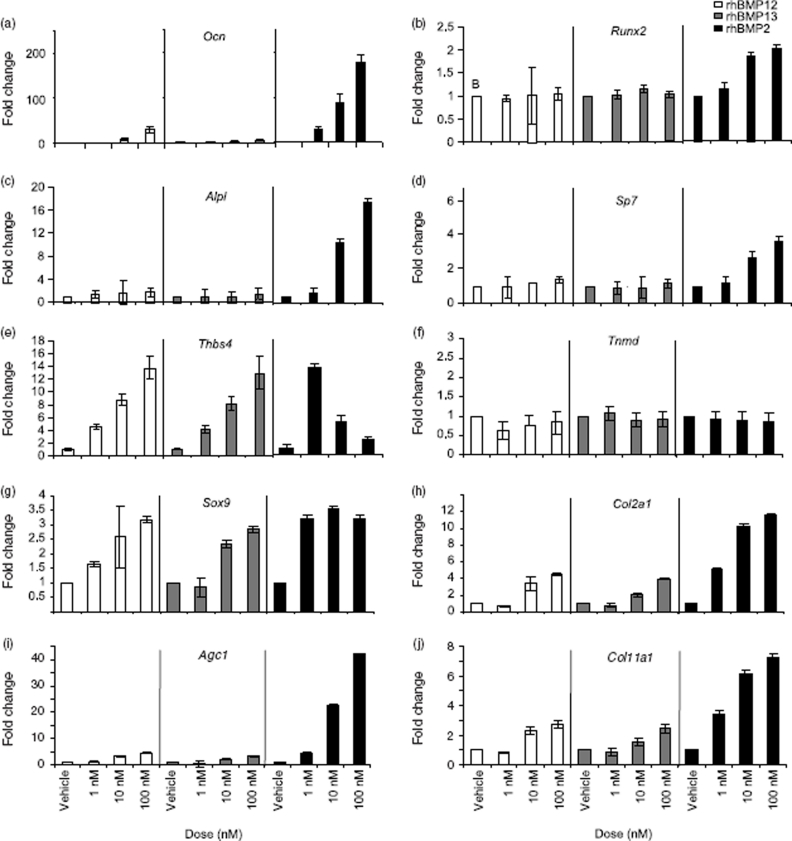
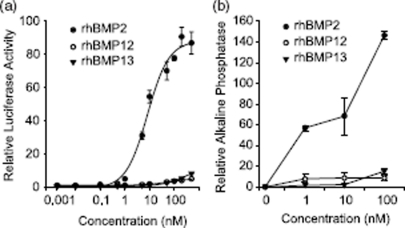
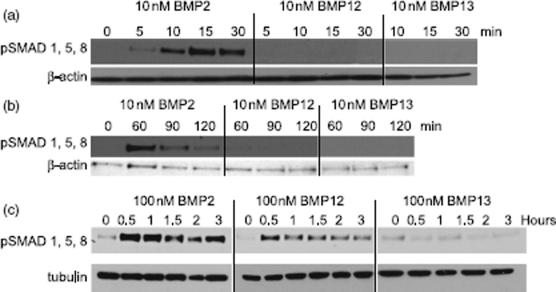
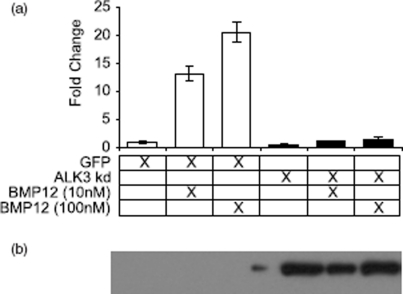
Ectopic expression of recombinant human bone morphogenetic protein 2 (rhBMP2) induces osteogenesis, while ectopic expression of rhBMP12 and rhBMP13 induces the formation of tendon-like tissue. Despite their different in vivo activities, all three ligands bound to the type I bone morphogenic protein receptors (BMPRs), activin receptor-like kinase (ALK)-3 and ALK6, and to the type II BMPRs, activin receptor type-2A, activin receptor type-2B, and BMPR2, with similar affinities. Treatment of C3H10T1/2 cells with rhBMP2 activated SMAD signaling and induced expression of osteoblast markers including osteocalcin mRNA (Ocn). In contrast, treatment with rhBMP12 or rhBMP13 resulted in a dose-dependent induction of a tendon-specific gene (Thbs4) expression with no detectable activation of SMAD 1, 5, and 8. Differential regulation of Thbs4 and Ocn has potential utility as an in vitro biomarker for induction of tenogenic signaling. Such an assay also permits the ability to distinguish between the activities of different BMPs and may prove useful in studies on the molecular mechanisms of BMP tenogenic activity.
Figures
References
-
- Amthor H, Christ B, Weil M, Patel K. The importance of timing differentiation during limb muscle development. Curr Biol. 1998;8:642–652. - PubMed
-
- Aspenberg P, Forslund C. Enhanced tendon healing with GDF 5 and 6. Acta Orthop Scand. 1999;70:51–54. - PubMed
-
- Bolt P, Clerk AN, Luu HH, Kang Q, Kummer JL, Deng ZL, Olson K, Primus F, Montag AG, He TC, Haydon RC, Toolan BC. BMP-14 gene therapy increases tendon tensile strength in a rat model of Achilles tendon injury. J Bone Joint Surg Am. 2007;89:1315–1320. - PubMed
-
- Chang SC, Hoang B, Thomas JT, Vukicevic S, Luyten FP, Ryba NJ, Kozak CA, Reddi AH, Moos M., Jr Cartilage-derived morphogenetic proteins. New members of the transforming growth factor-beta superfamily predominantly expressed in long bones during human embryonic development. J Biol Chem. 1994;269:28227–28234. - PubMed
-
- Cook SD, Rueger DC. Osteogenic protein-1: Biology and applications. Clin Orthop Relat Res. 1996:29–38. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous