

Genetic mechanisms involved in the evolution of the cephalopod camera eye revealed by transcriptomic and developmental studies
- PMID: 21702923
- PMCID: PMC3141435
- DOI: 10.1186/1471-2148-11-180
Genetic mechanisms involved in the evolution of the cephalopod camera eye revealed by transcriptomic and developmental studies
Abstract
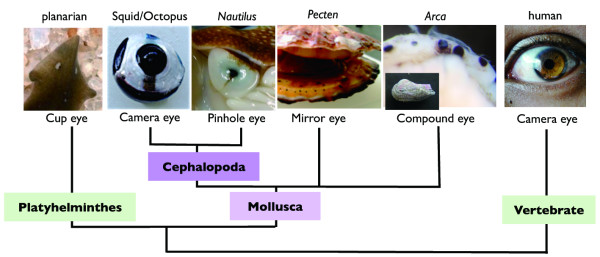
Background: Coleoid cephalopods (squids and octopuses) have evolved a camera eye, the structure of which is very similar to that found in vertebrates and which is considered a classic example of convergent evolution. Other molluscs, however, possess mirror, pin-hole, or compound eyes, all of which differ from the camera eye in the degree of complexity of the eye structures and neurons participating in the visual circuit. Therefore, genes expressed in the cephalopod eye after divergence from the common molluscan ancestor could be involved in eye evolution through association with the acquisition of new structural components. To clarify the genetic mechanisms that contributed to the evolution of the cephalopod camera eye, we applied comprehensive transcriptomic analysis and conducted developmental validation of candidate genes involved in coleoid cephalopod eye evolution.
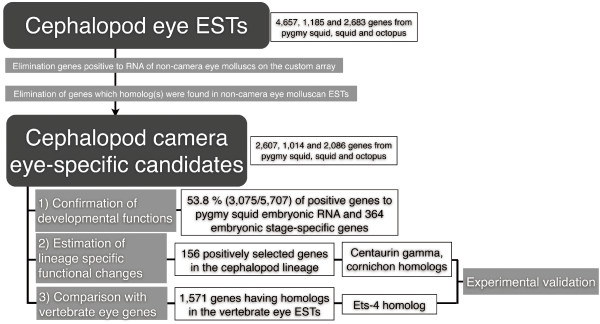
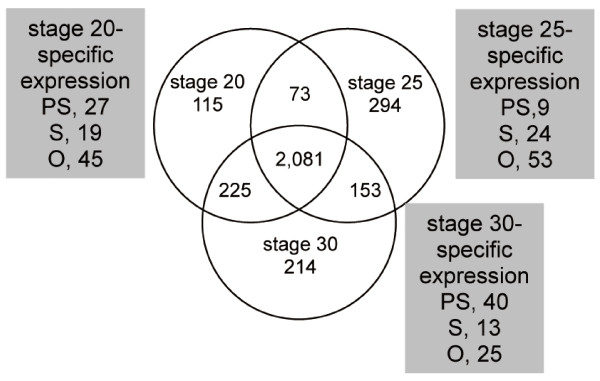
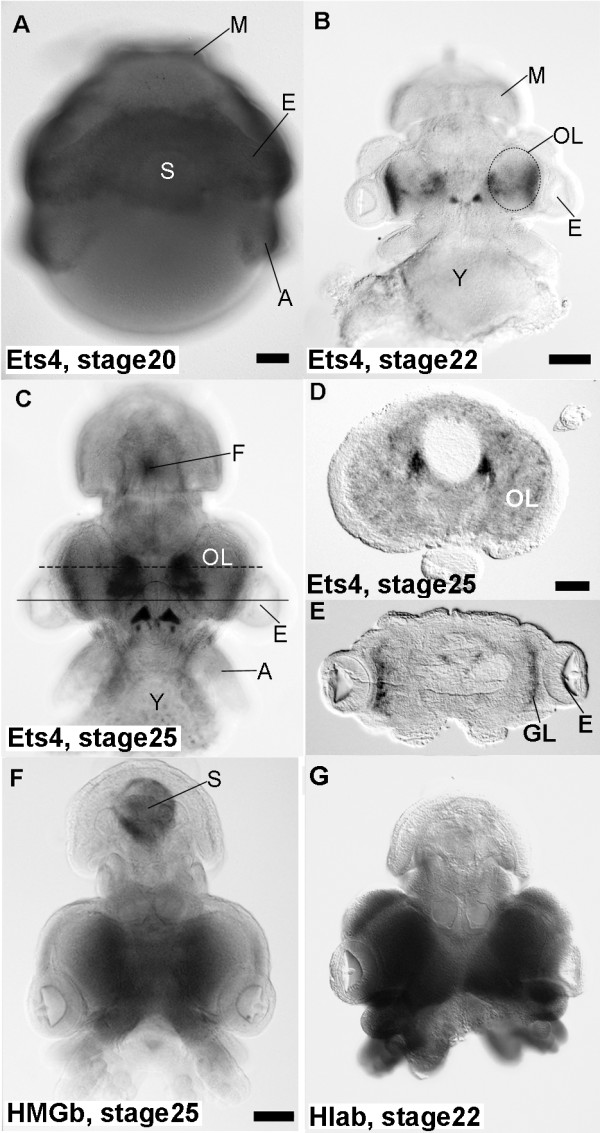
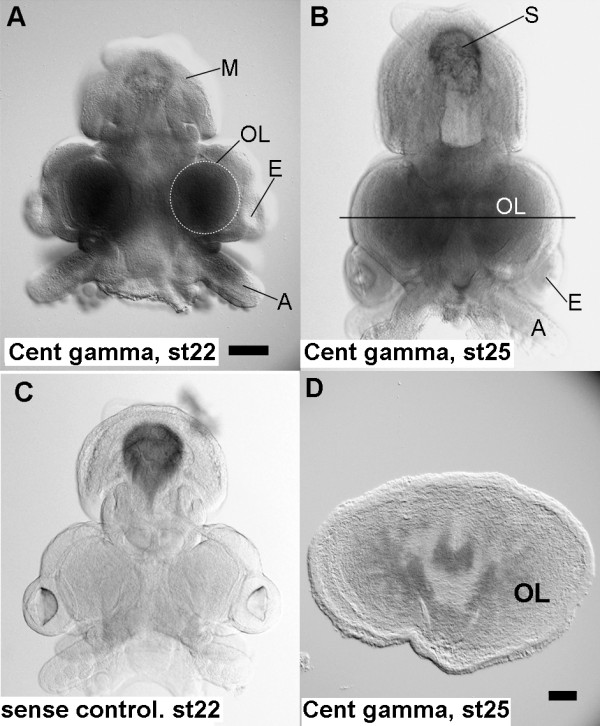
Results: We compared gene expression in the eyes of 6 molluscan (3 cephalopod and 3 non-cephalopod) species and selected 5,707 genes as cephalopod camera eye-specific candidate genes on the basis of homology searches against 3 molluscan species without camera eyes. First, we confirmed the expression of these 5,707 genes in the cephalopod camera eye formation processes by developmental array analysis. Second, using molecular evolutionary (dN/dS) analysis to detect positive selection in the cephalopod lineage, we identified 156 of these genes in which functions appeared to have changed after the divergence of cephalopods from the molluscan ancestor and which contributed to structural and functional diversification. Third, we selected 1,571 genes, expressed in the camera eyes of both cephalopods and vertebrates, which could have independently acquired a function related to eye development at the expression level. Finally, as experimental validation, we identified three functionally novel cephalopod camera eye genes related to optic lobe formation in cephalopods by in situ hybridization analysis of embryonic pygmy squid.
Conclusion: We identified 156 genes positively selected in the cephalopod lineage and 1,571 genes commonly found in the cephalopod and vertebrate camera eyes from the analysis of cephalopod camera eye specificity at the expression level. Experimental validation showed that the cephalopod camera eye-specific candidate genes include those expressed in the outer part of the optic lobes, which unique to coleoid cephalopods. The results of this study suggest that changes in gene expression and in the primary structure of proteins (through positive selection) from those in the common molluscan ancestor could have contributed, at least in part, to cephalopod camera eye acquisition.
Figures
Similar articles
-
Molecular Evidence for Convergence and Parallelism in Evolution of Complex Brains of Cephalopod Molluscs: Insights from Visual Systems.Integr Comp Biol. 2015 Dec;55(6):1070-83. doi: 10.1093/icb/icv049. Epub 2015 May 21. Integr Comp Biol. 2015. PMID: 26002349 Free PMC article.
-
Cephalopod eye evolution was modulated by the acquisition of Pax-6 splicing variants.Sci Rep. 2014 Mar 5;4:4256. doi: 10.1038/srep04256. Sci Rep. 2014. PMID: 24594543 Free PMC article.
-
Embryonic development of a centralised brain in coleoid cephalopods.Neural Dev. 2024 Jun 21;19(1):8. doi: 10.1186/s13064-024-00186-2. Neural Dev. 2024. PMID: 38907272 Free PMC article. Review.
-
Cephalopod versus vertebrate eyes.Curr Biol. 2023 Oct 23;33(20):R1100-R1105. doi: 10.1016/j.cub.2023.07.049. Curr Biol. 2023. PMID: 37875092
-
Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules: Extant cephalopods are younger than previously realised and were under major selection to become agile, shell-less predators.Bioessays. 2011 Aug;33(8):602-13. doi: 10.1002/bies.201100001. Epub 2011 Jun 17. Bioessays. 2011. PMID: 21681989 Review.
Cited by
-
A single-cell atlas of the bobtail squid visual and nervous system highlights molecular principles of convergent evolution.Nat Ecol Evol. 2025 Jul;9(7):1245-1262. doi: 10.1038/s41559-025-02720-9. Epub 2025 Jun 6. Nat Ecol Evol. 2025. PMID: 40481147 Free PMC article.
-
Krüppel-like factor/specificity protein evolution in the Spiralia and the implications for cephalopod visual system novelties.Proc Biol Sci. 2020 Oct 28;287(1937):20202055. doi: 10.1098/rspb.2020.2055. Epub 2020 Oct 21. Proc Biol Sci. 2020. PMID: 33081641 Free PMC article.
-
De novo assembly and characterization of two transcriptomes reveal multiple light-mediated functions in the scallop eye (Bivalvia: Pectinidae).PLoS One. 2013 Jul 29;8(7):e69852. doi: 10.1371/journal.pone.0069852. Print 2013. PLoS One. 2013. PMID: 23922823 Free PMC article.
-
Molecular Evidence for Convergence and Parallelism in Evolution of Complex Brains of Cephalopod Molluscs: Insights from Visual Systems.Integr Comp Biol. 2015 Dec;55(6):1070-83. doi: 10.1093/icb/icv049. Epub 2015 May 21. Integr Comp Biol. 2015. PMID: 26002349 Free PMC article.
-
Developmental principles: fact or fiction.ScientificWorldJournal. 2012;2012:980151. doi: 10.1100/2012/980151. Epub 2012 Feb 15. ScientificWorldJournal. 2012. PMID: 22489210 Free PMC article.
References
-
- Bonini NM, Bui QT, Gray-Board GL, Warrick JM. The Drosophila eyes absent gene directs ectopic eye formation in a pathway conserved between flies and vertebrates. Development. 1997;124:4819–4826. - PubMed
-
- Shen W, Mardon G. Ectopic eye development in Drosophila induced by directed dachshund expression. Development. 1997;124:45–52. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
