

Residual embryonic cells as precursors of a Barrett's-like metaplasia
- PMID: 21703447
- PMCID: PMC3125107
- DOI: 10.1016/j.cell.2011.05.026
Residual embryonic cells as precursors of a Barrett's-like metaplasia
Abstract
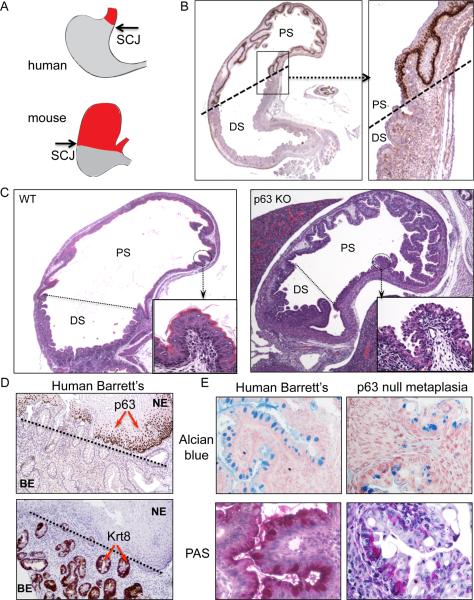
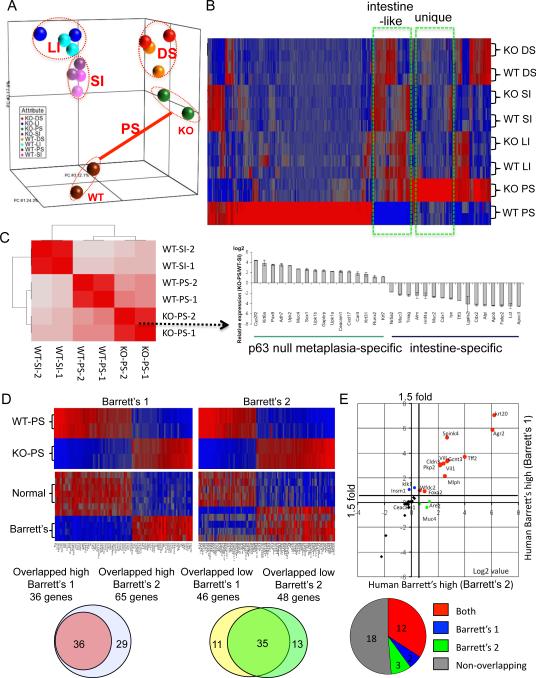
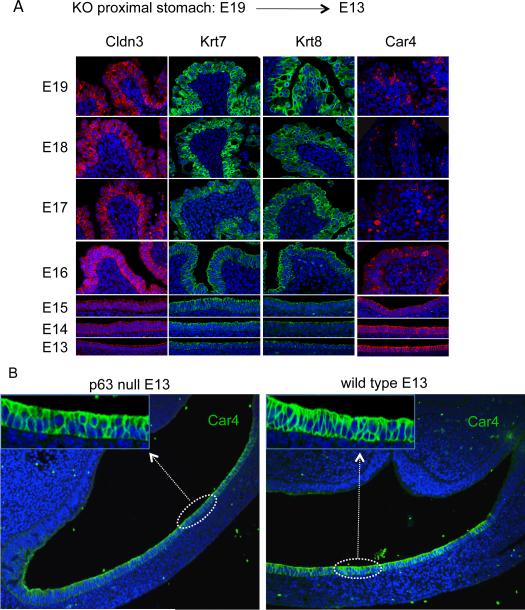
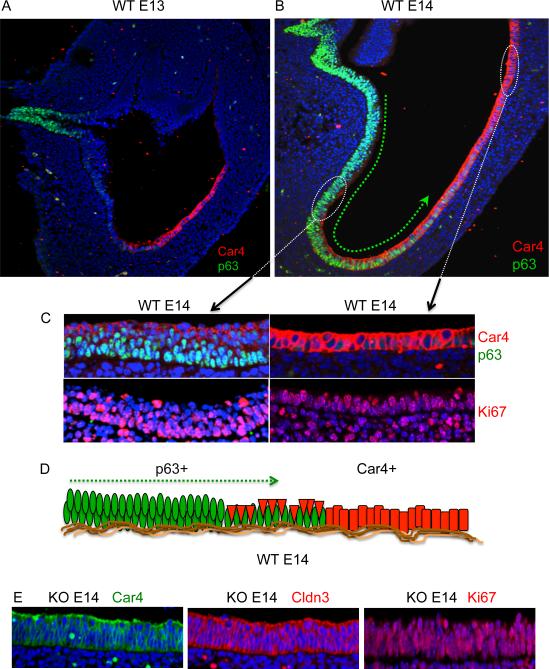
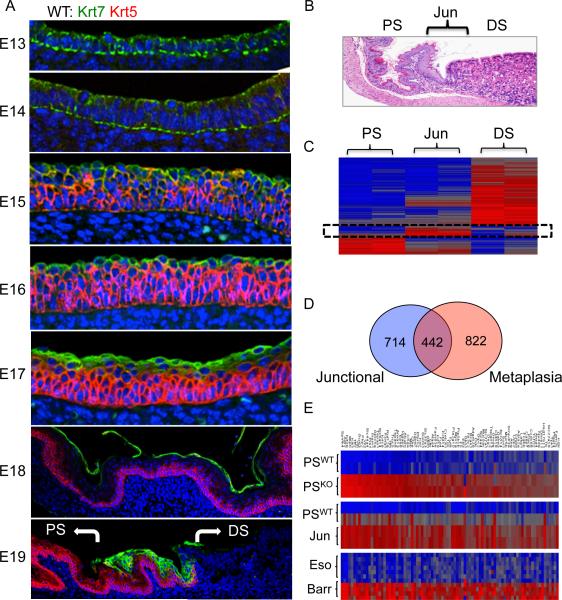
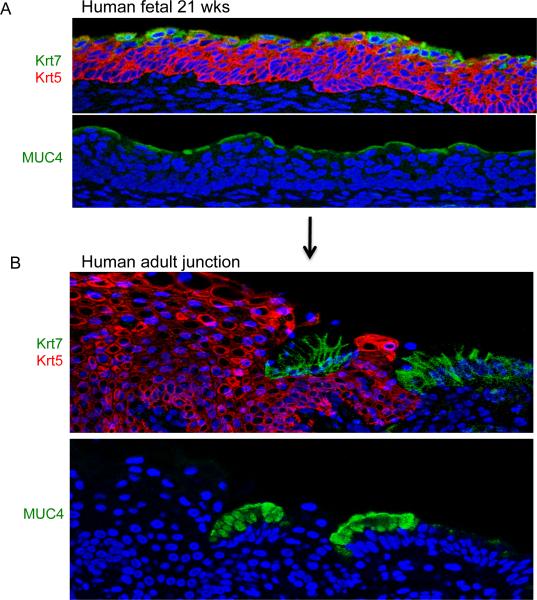
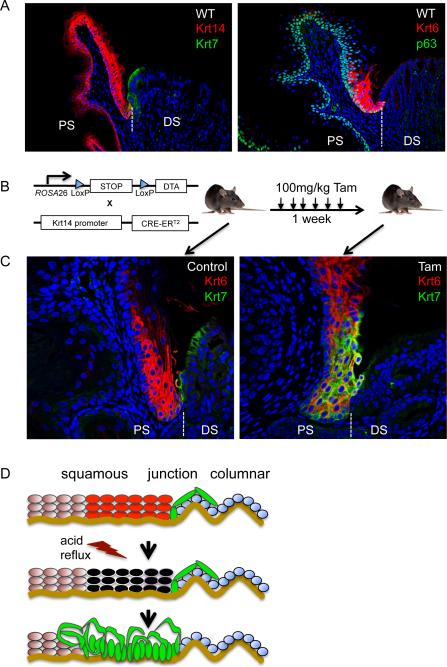
Barrett's esophagus is an intestine-like metaplasia and precursor of esophageal adenocarcinoma. Triggered by gastroesophageal reflux disease, the origin of this metaplasia remains unknown. p63-deficient mice, which lack squamous epithelia, may model acid-reflux damage. We show here that p63 null embryos rapidly develop intestine-like metaplasia with gene expression profiles similar to Barrett's metaplasia. We track its source to a unique embryonic epithelium that is normally undermined and replaced by p63-expressing cells. Significantly, we show that a discrete population of these embryonic cells persists in adult mice and humans at the squamocolumnar junction, the source of Barrett's metaplasia. We show that upon programmed damage to the squamous epithelium, these embryonic cells migrate toward adjacent, specialized squamous cells in a process that may recapitulate early Barrett's. Our findings suggest that certain precancerous lesions, such as Barrett's, initiate not from genetic alterations but from competitive interactions between cell lineages driven by opportunity.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
p63 and epithelial metaplasia: a gutsy choice.Cell. 2011 Jun 24;145(7):1003-5. doi: 10.1016/j.cell.2011.06.008. Cell. 2011. PMID: 21703442
-
Disease mechanisms: Out with the old, in with the new.Nat Rev Cancer. 2011 Jul 22;11(8):536. doi: 10.1038/nrc3116. Nat Rev Cancer. 2011. PMID: 21779007 No abstract available.
References
-
- Antonioli DA, Wang HH. Morphology of Barrett's esophagus and Barrett's-associated dysplasia and adenocarcinoma. Gastroenterol. Clin. North Am. 1997;26:495–506. - PubMed
-
- Badreddine RJ, Wang KK. Barrett's esophagus: an update. Nat. Rev. Gastroent. Hepatolff. 2010;7:369–378. - PubMed
-
- Barr H, Shepherd NA, Dix A, Roberts DJ, Tan WC, Krasner N. Eradication of high-grade dysplasia in columnar-lined (Barrett's) oesophagus by photodynamic therapy with endogenously generated protoporphyrin IX. Lancet. 1996;348:584–585. - PubMed
-
- Blot WJ, Devesa SS, Kneller RW, Fraumeni JF., Jr. Rising incidence of adenocarcinoma of the esophagus and gastric cardia. J. Am. Med. Assoc. 1991;265:1287–1289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
