

NLRX1 negatively regulates TLR-induced NF-κB signaling by targeting TRAF6 and IKK
- PMID: 21703539
- PMCID: PMC3150212
- DOI: 10.1016/j.immuni.2011.02.022
NLRX1 negatively regulates TLR-induced NF-κB signaling by targeting TRAF6 and IKK
Abstract
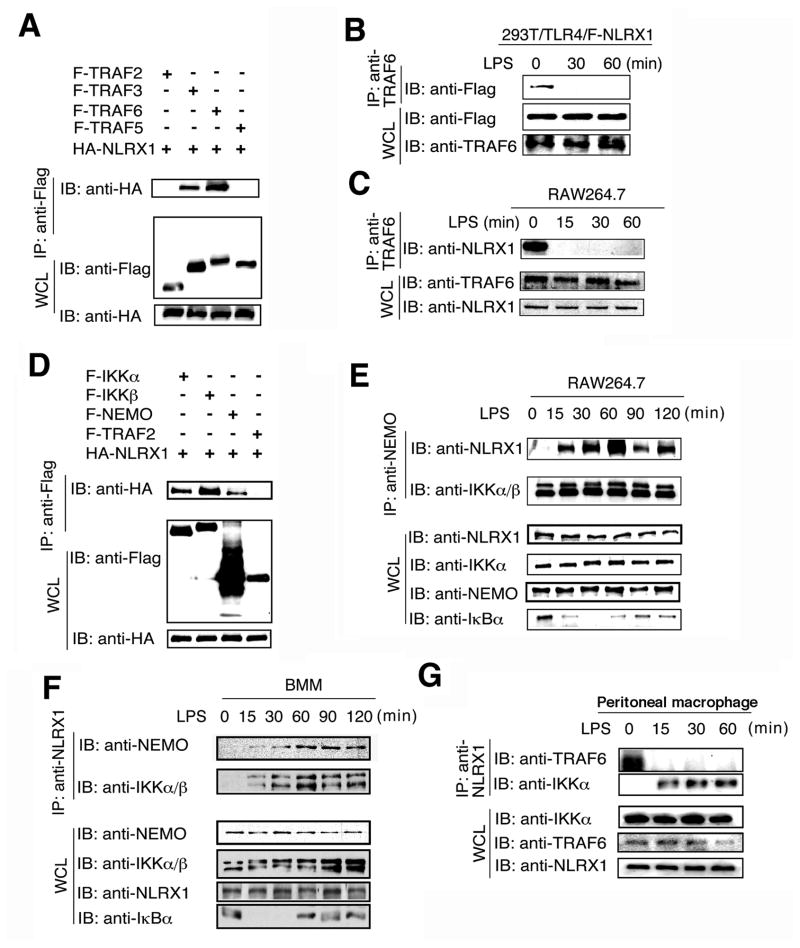
Tight regulation of NF-κB signaling is essential for innate and adaptive immune responses, yet the molecular mechanisms responsible for its negative regulation are not completely understood. Here, we report that NLRX1, a NOD-like receptor family member, negatively regulates Toll-like receptor-mediated NF-κB activation. NLRX1 interacts with TRAF6 or IκB kinase (IKK) in an activation signal-dependent fashion. Upon LPS stimulation, NLRX1 is rapidly ubiquitinated, disassociates from TRAF6, and then binds to the IKK complex, resulting in inhibition of IKKα and IKKβ phosphorylation and NF-κB activation. Knockdown of NLRX1 in various cell types markedly enhances IKK phosphorylation and the production of NF-κB-responsive cytokines after LPS stimulation. We further provide in vivo evidence that NLRX1 knockdown in mice markedly enhances susceptibility to LPS-induced septic shock and plasma IL-6 level. Our study identifies a previously unrecognized role for NLRX1 in the negative regulation of TLR-induced NF-κB activation by dynamically interacting with TRAF6 and the IKK complex.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures






Comment in
-
NOD so fast: NLRX1 puts the brake on inflammation.Immunity. 2011 Jun 24;34(6):821-2. doi: 10.1016/j.immuni.2011.06.006. Immunity. 2011. PMID: 21703534 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
