

Deficiency of the cytoskeletal protein SPECC1L leads to oblique facial clefting
- PMID: 21703590
- PMCID: PMC3135813
- DOI: 10.1016/j.ajhg.2011.05.023
Deficiency of the cytoskeletal protein SPECC1L leads to oblique facial clefting
Abstract
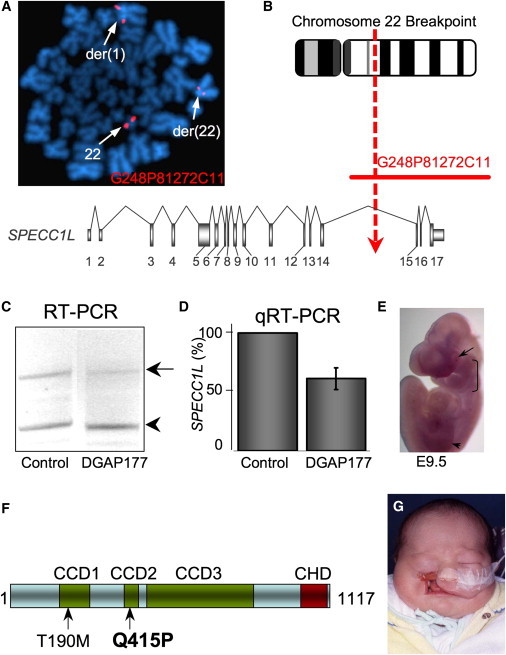
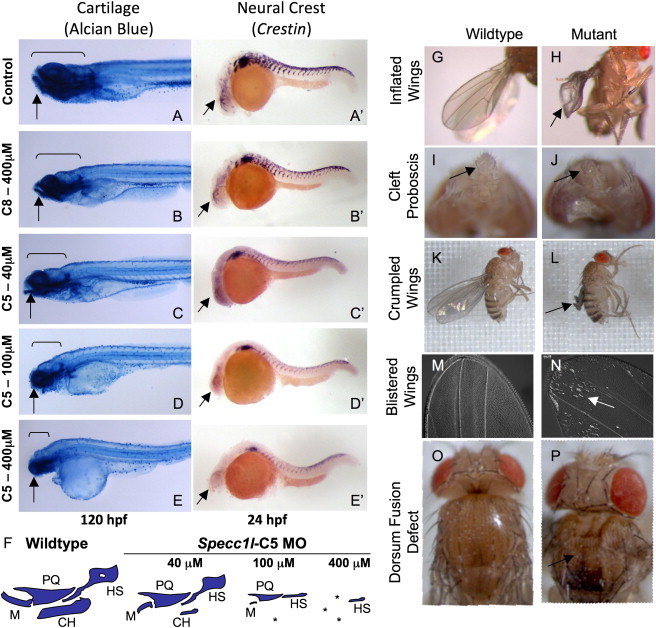
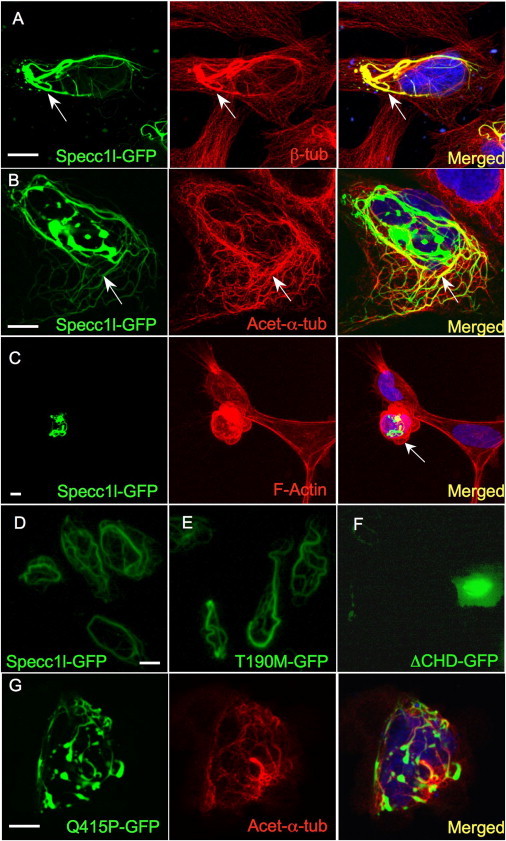
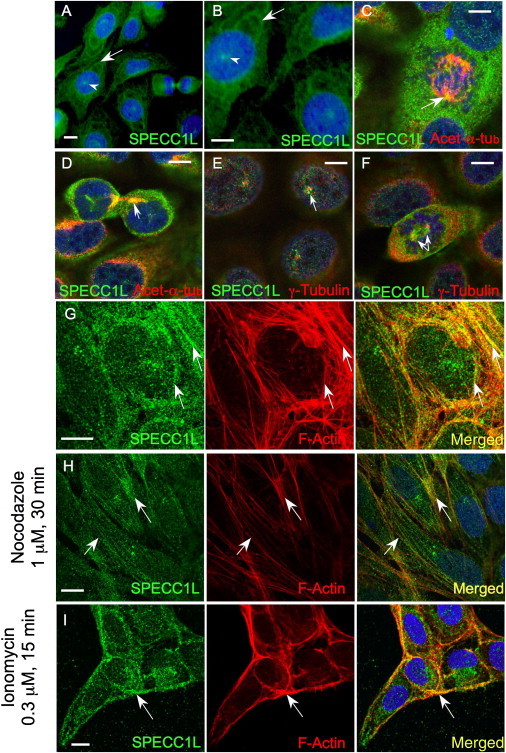
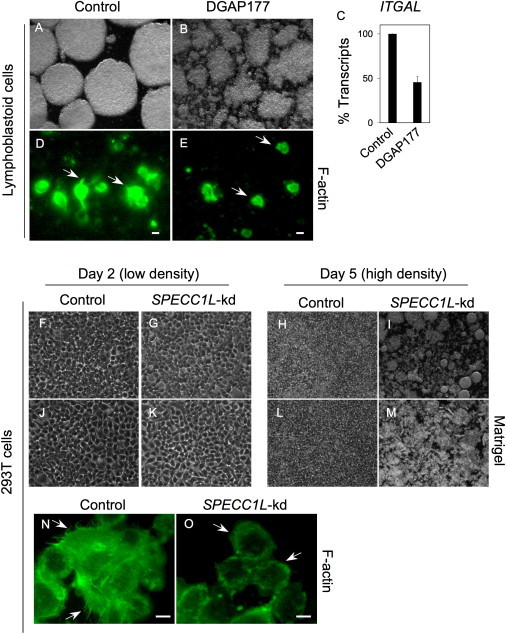
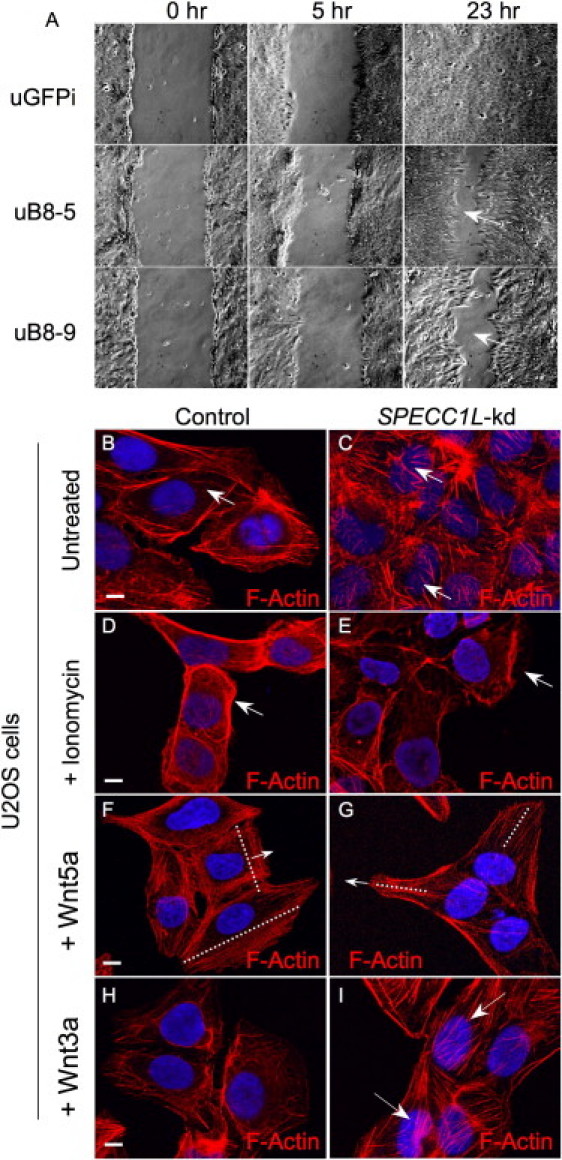
Genetic mutations responsible for oblique facial clefts (ObFC), a unique class of facial malformations, are largely unknown. We show that loss-of-function mutations in SPECC1L are pathogenic for this human developmental disorder and that SPECC1L is a critical organizer of vertebrate facial morphogenesis. During murine embryogenesis, Specc1l is expressed in cell populations of the developing facial primordial, which proliferate and fuse to form the face. In zebrafish, knockdown of a SPECC1L homolog produces a faceless phenotype with loss of jaw and facial structures, and knockdown in Drosophila phenocopies mutants in the integrin signaling pathway that exhibit cell-migration and -adhesion defects. Furthermore, in mammalian cells, SPECC1L colocalizes with both tubulin and actin, and its deficiency results in defective actin-cytoskeleton reorganization, as well as abnormal cell adhesion and migration. Collectively, these data demonstrate that SPECC1L functions in actin-cytoskeleton reorganization and is required for proper facial morphogenesis.
Copyright © 2011 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Jugessur A., Farlie P.G., Kilpatrick N. The genetics of isolated orofacial clefts: From genotypes to subphenotypes. Oral Dis. 2009;15:437–453. - PubMed
-
- Tessier P. Anatomical classification facial, cranio-facial and latero-facial clefts. J. Maxillofac. Surg. 1976;4:69–92. - PubMed
-
- Eppley B.L., van Aalst J.A., Robey A., Havlik R.J., Sadove A.M. The spectrum of orofacial clefting. Plast. Reconstr. Surg. 2005;115:101e–114e. - PubMed
-
- Hall B.K. Springer Science+Business Media; New York: 2009. The Neural Crest and Neural Crest Cells in Vertebrate Development and Evolution.
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
